

FULL RESEARCH ARTICLE
Robert M. Sullivan*
Northern California Wildlife Ecology and Fisheries Sciences, Weaverville, CA 96093, USA
*Corresponding Author: robert.m.sullivan@icloud.com; https://www.researchgate.net/profile/Robert-Sullivan-4
Published 22 November 2022 • www.doi.org/10.51492/cfwj.108.22
Abstract
I evaluated the impact and extent of the Monument Fire on the geographic range and suitable macrohabitat of the Trinity bristle snail (Monadenia setosa), a California endemic with limited distribution in northern California. Total area burned by the fire was ~87,984 ha or 46.0% of the species range (n = 191,156 ha). Total area of suitable macrohabitat for the species is ~107,913 ha of which 44.5% (n = 47,962 ha) was encompassed by the fire. Results show that the total area of forest cover-type vegetation and individual forest stand attributes impacted by the fire was not significantly different from areas within the species range not burned by the fire. There was no significant proportional differences in the six sequential categories of suitable macrohabitat burned by the fire (i.e., Low, Low-moderate, Moderate, Moderate-high, High, Critical suitability). The percentage of Moderate and Moderate-high suitable macrohabitat burned was only somewhat greater than predicted by the pre-fire species habitat suitability model (HSM). Many individual watersheds were encompassed by the fire and the resulting mosaic of burned watersheds was highly variable. Application of the Soil Burn Severity (SBS) map identified 8,293 ha (17.3%) of Unburned or very low burned soil, 24,191 ha (50.5%) of Low burned soil, 13,998 ha (29.2.1%) of Moderately burned soil, and 1,460 ha (3.0%) of Highly burned soil within the boundaries of the Monument Fire. When applied to categories of suitable macrohabitat, I calculated that 31,096 ha (100%) of Low to Low-moderate and 13,998 ha (96.1%) of Moderate to Moderate-high suitable macrohabitat were burned. High and Critical areas of macrohabitat suitability were much less impacted by high SBS (n = 1,461 ha [58.0%] because these regions were small in size, highly fragmented, widely dispersed across the landscape, and separated by major topographic and riverine discontinuities.
Key words: fuels reduction, landscape, macrohabitat, management, Monadenia setosa, terrestrial gastropod, threatened species, watershed
Introduction
Wildfires are a common natural occurrence in forest ecosystems of the Pacific Northwest (Burke et al. 1999). In northern California, decades of fire suppression, exclusion, and selective harvesting of timber has resulted in dense mixed-conifer and woodland forests, and overly congested riparian corridors. This condition is in large part a direct consequence of the lack of a comprehensive and coordinated program for long-term annual fuels reduction (Collins et al. 2019). Although natural fire regimes are often advocated, absence of progress in management of annual fuels reduction, at both state and federal levels, has greatly increased the susceptibility to fuels-driven mixed- and high-severity fire regimes (Wright and Agee 2004; Hessburg et al. 2005; Huff et al. 2005). Mixed- and high-severity fire regimes are potentially complex because of the combined influence of fuels variance, complex topography, stochastic climatic conditions, and fire suppression actions that produce intricate mosaic burn patterns (Halofsky et al. 2011). While climate has played a role in recent fire events and tree mortality, current forest conditions and lack of management are unquestionably contributing to both (Collins et al. 2019).
In recent decades the frequency and extent of fire has become one of the main ecological problems facing management and conservation planning of special status species, both in the context of prevention and restoration. The interaction between fire, fire management, and maintaining habitat for listed and rare species is of particular interest and importance within forests landscapes at high risk of fire (Agee 1993; Brown et al. 2004; Noss et al. 2006; Prather et al. 2008). As a consequence, the potential for high-severity forest fire has been identified as one of the primary risk factors for conservation of endemic species of terrestrial gastropods (Burke et al. 1999; Duncan et al. 2003; Duncan 2005). Numerous studies have documented that fire exerts a major impact on terrestrial snail communities by strongly reducing plant diversity and species richness (Nekola 2002; Kiss and Magnin 2006; Santos et al. 2009; Bros et al. 2011; Gaines et al. 2005, 2011). This is because wildfire-caused removal of vegetative cover and opening up the vegetation matrix fundamentally changes light and humidity levels, which are major threats to the survival of land snail populations. Still, there are a diversity of conclusions regarding the impact of wildfire on terrestrial gastropods (Whelan 1995; Caturla et al. 2000; Pausas and Verdu 2005; Moretti et al. 2009). For example, Bros et al. (2011) found that postfire management produced variation in habitat structure according to different levels of wood removal and the moist microhabitats provided by accumulated wood debris on the ground. In contrast, taxa occupying deep ground fissures may survive wildfire if associated with surface and buried rock accumulations, even though high intensity fires may eliminate most individuals in local populations, their microhabitat requirements, and primary food sources. As such, understanding mechanistic causes of this heterogeneous response is a conservation concern for the management of regional wildlife areas heavily affected by fire (Kiss and Magnin, 2002, 2003; Kiss et al. 2004; Santos et al. 2009; Driscoll et al. 2010).
Currently, there are ~113 species of terrestrial snails on the California Department of Fish and Wildlife Special Animal list (CDFW 2022; CNDDB 2022). The Trinity bristle snail (Monadenia setosa) is the only land snail in California with a threatened listing. (CDFW 1972). This species is endemic to watersheds of the Greater Trinity Basin, Klamath Bioregion, northern California (Talmadge 1952; Fig. 1). It inhabits older growth stands of mixed conifer and woodland forests, and riparian corridors of high decadence and dense understories. These conditions characteristically include standing deadwood and down logs important in substrate stability and juvenile survival (Roth 1978). Accumulations of live individuals are found most frequently in deep accumulations of surface or buried boulders with well-developed subterranean structure, and internal spacing within a stable saxicolous matrix, or in association with large underground root systems (Sullivan 2022b).

The recent Monument Fire was the largest wildfire to impact the Greater Trinity Basin in the last 20 years (Fig. 1). It was caused by a lightning strike in the vicinity of the town of Del Loma near Monument Peak, Trinity County on 16 August 2021. As of 18 September 2021, the area encompassed by the Monument Fire was estimated to be 87,984 ha or ~ 46.0% of the geographic range M. setosa. At a macroscale, this fire provided a unique opportunity to assess the potential impacts of wildfire on categories of suitable macrohabitat for the Trinity bristle snail (Sullivan 2022a). Lack of knowledge about the degree of coverage by the Monument Fire at the ground level precludes a discussion of the extent of microhabitat loss to the species without ground-truthing, given both the heterogeneity of the fire behavior and the mosaic distribution of polygons of suitable macrohabitat. However, the extent of microhabitat loss due to severe soil burn can be approximated at the macrohabitat level by superimposing a recent Soil Burn Severity map (SBS) onto the original habitat suitability model (HSM) for the species.
Irrespective of protective status, no study has ever evaluated the impact of a high-severity forest fire on an individual species or a community of terrestrial mollusks at a landscape-level in California. Land snails constitute an excellent model to study the impact of fire and the subsequent patterns of re-colonization, because they are highly sensitive to fire and have low dispersal abilities (Baur 1986; Baur and Baur 1990; Hansson 1991). Moreover, implementation of post-fire restoration and pre-treatment of local populations with low intensity fire management are actions easily surveyed and monitored. As such, my objectives were two-fold. First, I assess the impact of the Monument Fire on the geographic distribution of hypothesized suitable macrohabitat of the species using GIS-based fire and soil burn severity shapefiles applied to subsets of a recent habitat suitability model (HSM; Sullivan 2022a). Second, I examine the differential extent and severity of burning to categories of preferred suitable macrohabitat at both landscape and watershed levels. This information should prove useful to resource managers in understanding and evaluating the potential impact and response of this species to wildfire given its unique biology and ecology (Sullivan 2021, 2022a, 2022b). My study also provides a knowledge base for post-fire restoration, conservation, and biological diversity assessments of molluscoid forest-dwelling faunas in this fire prone region of northern California (Driscoll et al. 2010; Dunk et al. 2004).
Methods
Study Area
My study area was located in part of the Klamath Bioregion centered on the Greater Trinity Basin watershed, which includes geographic regions throughout the northwestern segment of the Trinity River and its tributaries in Trinity and Humboldt counties. The geographic range of M. setosa is ~191,156 ha (Fig. 1). This area is 25.6 % (745,817 ha) of the area encompassed by the Greater Trinity Basin, 20.3% (830,725 ha) of the area encompassed by Trinity County, but only 20.6% (927,728 ha) of the jurisdictional boundary of Humboldt County. This segment of the Klamath Bioregion extends to south-central Trinity County. This area is dominated by Klamath montane forest of Douglas fir (Pseudotsuga menziesii), white fir (Abies concolor), ponderosa pine (Pinus ponderosa), sugar pine (P. lambertiana), incense cedar (Calocedrus decurrens), tanoak (Lithocarpus densiflorus), and Pacific madrone (Arbutus menziesii) cover-types; and interdigitates with montane coastal forest of the North Coast Bioregion (Welsh 1994). Watersheds within this bioregion are almost entirely within mountains, with the only level land in a few narrow valleys (i.e., Weaverville Basin, Hyampom, and Hayfork valleys). These areas are dominated by mixed conifer and hardwood forest, riparian corridors of white alder (Alnus rhombifolia), big leaf maple (Acer macrophyllum), and various species of willow (Salix spp.). Upland environs are characterized by a deciduous hardwood understory of Pacific madrone, giant chinquapin (Castanopsis chrysophylla), tanoak, and canyon live oak (Quercus chrysolepis) cover-types.
Climate of this region is Mediterranean, with cool, wet winters and hot, dry summers. Annual precipitation over the Trinity River watershed averages ~1,400 mm. Precipitation ranges from 940 mm in lowlands (Weaverville and Hayfork) to as high as 2,200 mm (Barrett 1966). High rainfall combined with rugged geography results in extremely fast runoff and a high risk of flooding during winter storms. Soils are dominated by brown forest, gray-brown podzolic (i.e., acidic), and some immature soils, and Sierra-Nevada rock types (Mesozoic eugeosynclinal formations) intruded by Mesozoic granite (Welsh 1994). Large volumes of rocks and sediment carried by floods are spread along the rivers forming wide alluvial channels.
Data Collection
Field surveys focused on historical qualitative accounts of suitable habitat for the species based on documented occurrences (Talmadge 1952; Walton 1963; Roth 1978; Roth and Eng 1980; Sullivan 2022a). I sampled for active bristle snails during warm, wet, foggy, or rainy conditions during March to May and September to October 2008 and 2009. Typical microhabitat for this species included the surface of the soil, under objects large enough to accommodate a large (3.0–3.8 cm) adult shell (e.g., under boulders, slabs of thick sluffed-off bark from snags, dead wood, talus), on tree trunks and dead standing branches at the base of Pacific madrone and tan-oak root wads, and in other crevices with a well-developed organic soil base (Martin and Sommer 2004). I hand-picked snails in focal areas using the opportunistic visual search method, which was rapid (~30 min/per site) and entailed neither degradation nor soil removal (Gotmark et al. 2008; Raheem et al. 2008; Sullivan 2022b). Because land snails are dependent on microhabitat, different search images were required to prevent bias depending upon what substrate was encountered (i.e., boulder vs. tree vs. depression vs. flat ground; Fontaine et al. 2007; Cucherat and Demuynck 2008).
I derived model variables from descriptions of “suitable” habitat based on historical accounts for Monodenia spp. snails (Roth 1982; Roth and Pressley 1986; Duncan et al. 2003; Furnish et al. 1997). I used GIS-based landsat Visual Ecological Groupings (CALVEG; USFS 1981) and California Wildlife Habitat Relationships (CWHR; Airola 1988; Mayer and Laudenslayer 1988; Garrison et al. 2002) to assess geographic variation in forest type and stand structure (Parker and Matyas 1979; Goodchild et al. 1991; Sawyer and Keeler-Wolfe 1995).
A minimum mapping size of 2.5-ha pixels was used to contrast vegetation based on cover-type, vegetation type, tree cover from above, and over-story tree diameter. Non-forest abiotic environmental attributes were obtained from geo-rectified raster data sets for Northern California. Climate attributes were derived from the PRISM Climate Group (https://prism.oregonstate.edu/), and topographic variables (i.e., elevation and distance to nearest stream) were generated from 10-m digital elevation models (Table S1).
Partitioning the GIS-based Habitat Suitability Model
I used subsets of the original macrohabitat suitability model (hereafter referenced as “HSM”; Fig. 2) developed for M. setosa (Sullivan 2022a) to evaluate the impact of the Monument Fire on the geographic range of the species and affected watersheds. This HSM included collection of macrohabitat attributes at 333 sample sites, which were attached to 80,000 random points that encompassed the total range of the species. For comparative purposes, I partitioned the original HSM into areas encompassed by the Monument Fire (referenced as “Monument Fire HSM”; n = 47,962 ha) and areas not impacted by the fire (referenced as “Non-fire HSM”; n = 59,951 ha). This process allowed a comparison between fire- and non-fire impacted models for all environmental variables, categories of macrohabitat suitability, and SBS. The original HSM was developed using pre-fire CALVEG forest vegetation cover-type layers in combination with digital orthoquads of other environmental base-layers (ArcMap; ESRI 2022).

These variables were used as selection criteria in a GIS-based landscape-level HSM for the species to query vector-based polygons (shapefiles; Query Builder tool) throughout the range of the species. I used point density surfaces, parameterized by theoretical point-density functions representing hypothetical limits, which defined a range of relative macrohabitat suitability. I then compared point density surfaces among HSM groups and burned watersheds. Six categories of macrohabitat suitability were identified: Low, Low-moderate, Moderate, Moderate-high, High, and Critical.
Soil Burn Severity
Raw SBS data (raster scale = 30 m) were converted to a polygon (vector) shapefile to assess the mosaic pattern of SBS in the Monument Fire-HSM (Raster-to-Polygon tool, ArcMap). These data were provided by the Burned Area Emergency Response Program (BAER 2022) and the United States Forest Service, Weaverville District (K. Bloomer, pers. comm.), and involved use of remote sensing imagery with field validated soils data to develop the final “best” fit SBS map. I approximated the degree of coverage at ground level by overlaying the SBS map onto the shapefile of suitable macrohabitat encompassed by the Monument fire. Categories of SBS in the layer included four levels: 1) Unburned-very low, 2) Low, 3) Moderate, and 4) High. Low SBS indicates only partial consumption of fine fuels while litter coverage remains relatively intact on the soil surface.
Moderate SBS indicates consumption and conversion to ash of nearly all soil cover and vegetative litter and fine fuels. High SBS indicates high intensity fire behavior coincident with longer burn time at the surface of the soil. Due to high heat, nearly all soil covered by vegetative litter and fuels is consumed leaving bare soil prone to the impacts of precipitation and resulting water runoff. Importantly, the SBS model only characterizes the effects of wildfire on the soil surface and below-ground. It does not assess the effects of mortality to vegetation or canopy alteration (Abele 2010). Changes in soil cover, water repellency, and soil physical and biological changes determine the level of severity in burnt soil. These environmental elements are critical to survival and viability of populations of terrestrial gastropods and other soil-dwelling invertebrates important in maintaining soil quality (Martin and Sommer 2004).
Statistical Analyses
I performed all analyses using R Statistical Software (R Core Team 2022) and set statistical significance at α < 0.05. I evaluated univariate normality of all GIS-based environmental variables by use of the adjusted Anderson-Darling (AD.adj) statistic (Razali and Wah 2011). All tests for normality failed for both the fire HSM (AD = < 1,801.0 and > 55.0, P < 0.001, n = 4,668) and the non-fire HSM (AD = < 2,698.0 and > 86.0, P < 0.001, n = 6,971). Thus, I used nonparametric statistical tests to evaluate significance between each HSM for all environmental parameters. I used the Kruskal-Wallis non-parametric analysis of variance (ANOVA) rank sum test (ꭓ2) to evaluate the null hypothesis of no significant difference between subsets of the original HSM for each environmental parameter. I used Spearman’s rank correlation (rs; 2-tailed test) to calculate the strength and direction of the relationship between any two variables whether linear or not (Corder and Foreman 2014). I used paired samples Wilcoxon signed rank (V) test (Rosner et al. 2006) to compare the proportion of combined perimeter (m) and ha metrics among categories of suitable macrohabitat between both partitions of the original HSM.
Because environmental data were not normally distributed, I used Generalized Linear Modeling (GLM; family = Gamma [link = “log”]; Nelder and Wedderburn 1972) to evaluate the relationship between each environmental variable and the six categories of macrohabitat suitability for both partitions of the original HSM. I used a Poisson error-structure (family = Poisson [link = “log”]) with each GLM regression when comparing habitat affected by categories of both habitat suitability and SBS to graphically establish the relationship between the response variables (i.e., suitability and SBS) and smoothed functions of the predictor variable (i.e., ha of habitat). I used this error structure because data consisted of categories (i.e., suitability and SBS) and counts (ha). I used the residual deviance statistic (ꭓ2) to evaluate the difference between null and residual deviances to evaluate if the model was “useful” in fitting the data in each environmental subset assessed by GLM (i.e., pchisq = null deviance – residual deviance, with degrees of freedom [df] = number of predictor variables in the model (https://www.statology.org/null-residual-deviance).
I constructed plots of the relationships among the six categories of macrohabitat suitability using linear discriminant function (DFA) analyses on log-transformed environmental variables for those attributes found to be significantly correlated with suitability criteria based on GLM analyses. DFA uses linear combinations of the predictors to create new axes for the final classification. DFA strives to find the linear combinations of the original environmental variable subsets (i.e., forest stand, monthly minimum and maximum temperature, precipitation) that gives the best possible separation between subsets of the original HSM dataset. This process maximizes component axes for class-separation and provides a visual assessment in 2-dimensional space of major variable-driven differences between each category of suitability for both partitions of the original HSM. I used confusion statistics (ConfusionMatrix function, Program “caret”; Accuracy and Kappa values) to assess the “fit” of the DFA model for each category of habitat suitability. Accuracy refers to the “success” rate of the model expressed as a percent. The Kappa statistic is a normalized statistic that compares the accuracy of the observed model to the accuracy of an expected random model; its value never exceeds 1.0.
Results
Comparison of Environmental Variables between Subsets of the HSM
The primary concentration of the Monument Fire occurred in the northeast section of the species range on both sides of the Trinity River, which included the southernmost sectors of tributaries of the North Fork Trinity River and New River near the confluence with the mainstem Trinity River. My analysis showed that the Monument Fire encompassed 44.5% (47,962 ha) of the total available area of suitable macrohabitat for the species (~107,913 ha). A comparison of the total ha within the CWHR cover-type classification found similar patterns in the proportion of each vegetation type between the Monument Fire and Non-fire partitions of the original HSM (Table 1). For each subset of the HSM the predominant vegetation type was Douglas fir forest followed by montane hardwood conifer, and Sierra mixed conifer, which was slightly more prevalent in the Monument Fire HSM compared to the Non-fire HSM (Table 2). Blue oak-foothill pine and montane hardwood were virtually nonexistent for each subgroup of the original HSM.
Table 1. Standard statistics based on ha of various vegetation types contained within the California Wildlife Habitat Relationship System (CWHR) cover-type Geographic Information System (GIS) layer for each subset of the habitat suitability model (HSM for the Monument Fire (MF) and the Non-fire (NF) HSMs.
Table 1a. Monument Fire HSM
| Vegetation type | Polygon count | Min. | Max. | x̄ | n | % | SD | S2 |
|---|---|---|---|---|---|---|---|---|
| Blue oak-foothill pine | 52 | 0.1 | 14.3 | 1.5 | 77 | 0.7% | 2.2 | 4.7 |
| Douglas fir | 2,131 | 0.0 | 57.3 | 2.8 | 5,912 | 55.1% | 4.1 | 16.7 |
| Montane hardwood conifer | 1,783 | 0.0 | 38.5 | 1.8 | 3,137 | 29.2% | 2.3 | 5.3 |
| Montane hardwood woodland | 23 | 0.7 | 8.7 | 2.6 | 60 | 0.6% | 2.2 | 4.9 |
| Sierra mixed conifer | 679 | 0.0 | 21.5 | 2.3 | 1,541 | 14.4% | 2.8 | 7.5 |
| Total | 4,668 | — | — | — | 10,728.70 | 100% | — | — |
Table 1b. Non-fire HSM
| Vegetation type | Polygon count | Min. | Max. | x̄ | n | % | SD | S2 |
|---|---|---|---|---|---|---|---|---|
| Blue oak-foothill pine | 50 | 0.0 | 5.2 | 1.1 | 53 | 0.4% | 1.0 | 0.9 |
| Douglas fir | 3,664 | 0.0 | 46.0 | 2.1 | 7,820 | 60.0% | 3.0 | 9.2 |
| Montane hardwood conifer | 2,642 | 0.0 | 25.8 | 1.5 | 4,042 | 31.0% | 1.8 | 3.1 |
| Montane hardwood woodland | 13 | 0.4 | 8.6 | 2.1 | 28 | 0.2% | 2.1 | 4.4 |
| Sierra mixed conifer | 602 | 0.0 | 16.1 | 1.8 | 1,100 | 8.4% | 2.0 | 3.9 |
| Total | 6,971 | — | — | — | 13,042 | 100% | — | — |
Table 2. Perimeter and area of suitable habitat categories for the Monument Fire (MF) and the Non-fire (NF) HSMs.
| Suitability | MF Perimeter | MF Perimeter % | MF | MF ha % | NF Perimeter | NF Perimeter % | NF ha | NF ha % |
|---|---|---|---|---|---|---|---|---|
| Low | 1,250,610 | 34.8% | 18,576 | 38.7% | 1,789,364 | 40.6% | 27,209 | 45.4% |
| Low-moderate | 1,001,335 | 27.9% | 12,668 | 26.4% | 1,337,894 | 30.4% | 18,508 | 30.9% |
| Moderate | 747,140 | 20.8% | 9,100 | 19.0% | 796,398 | 18.1% | 9,312 | 15.5% |
| Moderate-high | 403,232 | 11.2% | 5,469 | 11.4% | 324,685 | 7.4% | 3,513 | 5.9% |
| High | 154,146 | 4.3% | 1,702 | 3.5% | 96,437 | 2.2% | 869 | 1.4% |
| Critical | 38,254 | 1.1% | 448 | 0.9% | 59,098 | 1.3% | 539 | 0.9% |
| Total | 3,594,716 | — | 47,962 | — | 4,403,876 | — | 59,951 | — |
Results of the Kruskal-Wallis non-parametric ANOVA showed that 73.9% (n = 46) of all environmental attributes were significantly different between the two HSM subsets (Table 3). All metrics describing forest stand elements within the boundaries of the Monument Fire HSM had significantly larger values compared to the Non-fire HSM. Similarly, except for averaged monthly minimum temperatures in March and April, all other values were significantly warmer for the Monument Fire HSM compared to the Non-fire HSM. This pattern was also evident for the months of June, July, September, and October for averaged maximum monthly temperatures. For all other months averaged maximum temperatures were warmer within the Non-fire HSM. In average monthly precipitation the Monument Fire-HSM received significantly less precipitation in every comparison that was significant with the Non-fire HSM.
Table 3. Summary of mean values for each GIS macrohabitat variable and results of the Kruskal-Willis (KW) nonparametric analyses (df = 2) of variance for the Monument Fire HSM (n = 4,668) versus the Non-fire HSM (n = 6,971). Variables not significant different between subsets are not shown. CFA = cover from above, OTD = over-story tree diameter, DBH = diameter breast height; P = 0.5*, P = 0.01**, P = 0.001***.
Table 3a. Forest stand, stream, and elevation variables
| Variables | Monument Fire HSM x̄ | Non-fire HSM x̄ | ꭓ2 | P-value |
|---|---|---|---|---|
| 1.Total tree CFA | 78.4 | 76.3 | 70.0 | 0.001*** |
| 2. Conifer CFA | 40.7 | 39.8 | 18.0 | 0.001*** |
| 3. Hardwood CFA | 32.7 | 31.5 | 45.0 | 0.001*** |
| 4.OTD (DBH) | 21.6 | 22.1 | 16.0 | 0.001*** |
| 5. CWHR size | 4.0 | 3.9 | 36.0 | 0.001*** |
| 6. Elevation | 803 | 794.2 | 10.0 | 0.01*** |
Table 3b. Average monthly minimum temperature (oC)
| Variables | Monument Fire HSM x̄ | Non-fire HSM x̄ | ꭓ2 | P-value |
|---|---|---|---|---|
| 11. March | 0.3 | 0.3 | 12.0 | 0.001*** |
| 12. April | 1.5 | 1.5 | 12.0 | 0.001*** |
| 13. May | 4.7 | 4.6 | 97.0 | 0.001*** |
| 14. June | 8.0 | 7.9 | 155.0 | 0.001*** |
| 15. July | 11.3 | 11.0 | 310.0 | 0.001*** |
| 16. August | 10.7 | 10.3 | 323.0 | 0.001*** |
| 17. September | 8.3 | 8.1 | 72.0 | 0.001*** |
| 18. Annual total | 4.0 | 3.9 | 69.0 | 0.001*** |
Table 3c. Average monthly maximum temperature (ºC)
| Variables | Monument Fire HSM x̄ | Non-fire HSM x̄ | ꭓ2 | P-value |
|---|---|---|---|---|
| 22. January | 8.3 | 8.5 | 293.0 | 0.001*** |
| 24. March | 13.8 | 13.9 | 11.0 | 0.001*** |
| 25. April | 17.2 | 17.3 | 14.0 | 0.001*** |
| 26. May | 22.3 | 22.4 | 6.0 | 0.050* |
| 27. June | 27.5 | 27.5 | 7.0 | 0.020* |
| 28. July | 32.7 | 32.6 | 40.0 | 0.001*** |
| 30. September | 29.2 | 29.1 | 52.0 | 0.001*** |
| 31. October | 22.1 | 21.9 | 129.0 | 0.001*** |
| 32. November | 12.2 | 12.4 | 231.0 | 0.001*** |
| 33. December | 8.2 | 8.4 | 366.0 | 0.001*** |
Table 3d. Average monthly precipitation (cm) and total annual evapotranspiration.
| Variables | Monument Fire HSM x̄ | Non-fire HSM x̄ | ꭓ2 | P-value |
|---|---|---|---|---|
| 35. January | 199.4 | 219.6 | 1,563.0 | 0.001*** |
| 36. February | 161 | 178.9 | 1,791.0 | 0.001*** |
| 37. March | 151.5 | 169.5 | 1,844.0 | 0.001*** |
| 38. April | 66.8 | 74.2 | 1,056.0 | 0.001*** |
| 39. May | 33.7 | 36.7 | 699.0 | 0.001*** |
| 40. June | 17.8 | 18.0 | 20.0 | 0.001*** |
| 43. October | 78.1 | 82.7 | 465.0 | 0.001*** |
| 44.November | 184.0 | 202.8 | 1,328.0 | 0.001*** |
| 45. December | 208.2 | 228.6 | 1,394.0 | 0.001*** |
| 46. Evapotranspiration | 1,147.6 | 1,257.5 | 1,353.0 | 0.001*** |
Comparison of Categories of Suitable Habitat between Subsets of the HSM
In both perimeter and area, the Monument Fire HSM had a somewhat smaller proportion of Low and Low-moderate suitability habitat, but a somewhat larger proportion of Moderate and High suitability habitat than the Non-fire HSM (Table 1; Fig. 3A-3D). Paired Wilcoxon signed rank exact tests showed no significant difference between HSMs in the proportion of suitable habitat categories when measurements of polygon perimeters and ha of habitat were combined in the paired comparison (V = 34.0, P = 0.965, n = 12); whereas pairwise correlations between categories were highly significant (rs = 0.980, P < 0.001, n = 12) .
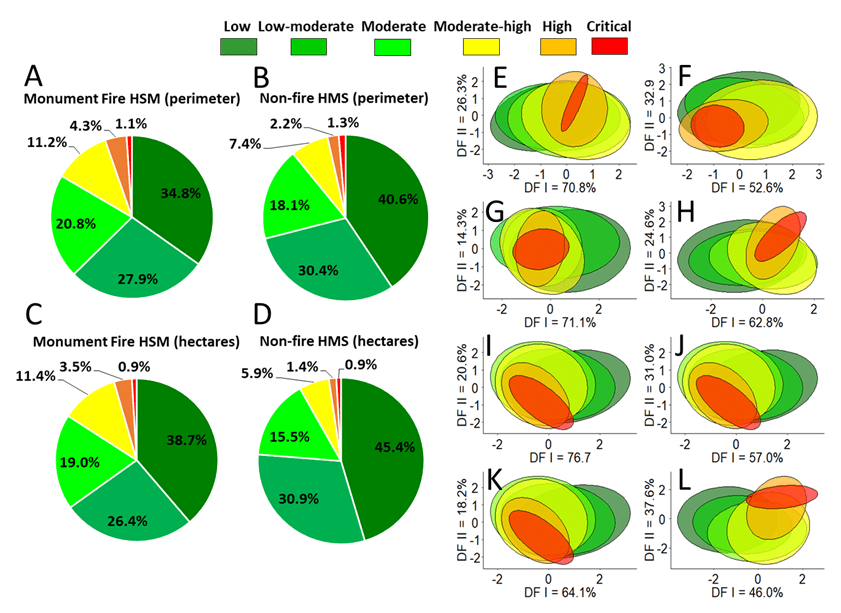
Comparisons among Categories of Habitat Suitability in Each Subset of the HSM
Mean values for High and Critical suitability values for each subcomponent of the original HSM were generally larger or smaller than any of the other macrohabitat suitability categories (Table S2). Among forest stand site attributes, GLM identified significant correlations between categories of suitability and total tree CFA, overstory tree diameter, WHR size, elevation, evapotranspiration, and distance to the nearest stream (Table S3). Average monthly minimum temperature suitability was significantly correlated with the months of January and February, April through June, and September through annual averaged monthly values. Categories of suitability were significantly correlated with averaged monthly maximum temperature for the months of January and February, May through July, and October through annual averaged monthly values. For averaged monthly precipitation all months were significantly correlated with suitability, annual averaged values. In all GLM regressions the residual deviance statistic (i.e., ꭓ2-P-value test) was significant, indicating that each regression model was “useful” in fitting the data in each subset of environmental attributes.
For both the Monument Fire and Non-Fire HSMs, forest stand characteristic and maximum monthly temperature explained the greatest amount of variation along DF I (70.8% and 76.7% versus 71.1% and 64.1%, respectively; Table S4). Along DF II the greatest amount of variation explained in the Fire HSM was for average minimum monthly temperature (32.9%) and forest stand characteristics (26.3%). For the Non-fire HSM it was average monthly precipitation (37.6%) and minimum monthly temperature (31.0%). Evaluation of the coefficients of linear discriminants (i.e., variable loadings) for DF I showed that for the Monument Fire HSM monthly: 1) evapotranspiration, 2) August, September, and annual minimum temperatures, 3) April maximum temperature; and 4) November precipitation had the strongest positive loadings on this discriminant vector, respectively. For the Non-fire HSM evaluation of variable loadings for DF I showed that: 1) elevation and average monthly, 2) annual minimum temperature, 3) maximum June temperature, and 4) November precipitation had the strongest positive loadings on DF I, respectively).
Subsets of environmental variables that were significantly correlated with suitability in the GLM regression analyses were used as input into the DFA plots of habitat suitability. Plots of the first two discriminate functions for both the Monument Fire HMS (Fig. 3E-3H) and Non-fire HMS (Fig. 3I-3L) showed that High and Critical macrohabitat suitability exhibited the least amount of variation along DF I and DF II. The extent of separation among categories of suitability graded from moderately negative to moderately positive along DF I for both subsets. Inspection of individual coefficients of linear discriminants and discriminant function plots showed that no environmental attribute definitively separated High and Critical from other categories of suitability at the 95% confidence level.
The greatest extent of overlap in variance occurred in Low, Moderate-low, and Moderate suitability, which encompassed the largest total acreages among categories of suitability for each HSM subset. Based on the 2-dimensional DF plots, the greatest separation between categories of both High and Critical suitability from other categories appeared to be in averaged monthly precipitation (Fig. 3H and 3L). For example, in the Fire HSM DF I (62.8%) was dominated by a positive November vs. negative September precipitation ratio; and DF II (34.6%) was dominated by a positive January versus a negative February-October precipitation ratio. For the Non-fire HSM DF I (46.0%) was dominated by a positive November vs. a negative annual-September precipitation ratio; and DF II (37.6%) was dominated by a positive December-September-April vs. a negative January-February precipitation ratio. This interpretation was also substantiated by the observation that the average monthly precipitation subset had somewhat higher Accuracy and Kappa statistics for both the Monument Fire (Accuracy = 36.4%, Kappa = 0.17) and Non-Fire HSMs (Accuracy = and 40.2%, Kappa = 0.17) compared to other environmental data.
Soil Burn Severity
Of the 90,195 ha of the Monument Fire evaluated in the SBS model 19.7% (n = 17,765) had Unburned-very low SBS, 47.1% (n = 42,472) had Low SBS, 29.1% (n = 26,260) had Moderate SBS, and 4.1% (n = 3,662) had High SBS. These proportions were virtually identical to those found in the 47,942 ha of the SBS map that encompassed the Monument Fire HSM (Fig. 4). Unexpectedly in terms of total ha of habitat, my analysis showed that each SBS category had virtually the same proportional degree of soil burn on each category of suitability in the Monument Fire HSM (Fig. 5A-5D).
Watersheds Encompassed by the Fire and Impacted by the Soil Burn Severity
There were 48 watersheds burned by the Monument Fire-HSM (Fig. 4). Of the 81,115 ha contained within the boundaries of these watersheds 59.3% (n = 46,071) were affected by the fire to some degree (Table S5). Minors, Canadian, Lower Big French, Don Juan, Middle Big French, Bear, Big creeks, and Dinner Gulch watershed had the largest concentrations of High and Critical suitable habitat for the Trinity bristle snail. Whereas, Rowdy Bar, Italian (type locality of M. setosa), Mill 1, Manzanita, and Dutch creek watersheds had Moderate-high habitat suitability. Although not every watershed contained large acreages of suitable habitat the relationship between total ha in the watershed and total ha of suitable habitat was significant (rs = 0.820, P < 0.001, n = 48).

In general, the bigger the watershed the more likely it contained suitable habitat for the species. However, these significant relationships only held for Low to Moderate-high categories because in most watersheds there were few or zero ha of High or Critical suitable habitat even in the largest of watersheds (Fig. 5E). Also, each category of suitable habitat graded into each adjacent category in terms of total ha. For example, Low suitability was significantly correlated with Low-moderate suitability, Low-moderate suitability was significantly correlated with Moderate suitability, Moderate suitability was significantly correlated with Moderate-high suitability, Moderate-high suitability was significantly correlated with High and Critical suitability, and High suitability was significantly correlated with Critical suitability (Table 4).
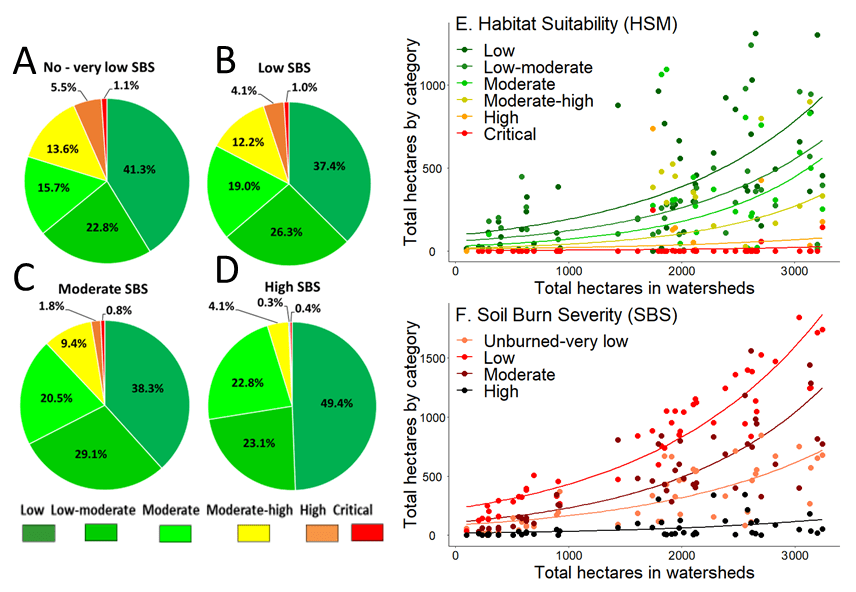
Table 4. Spearman rank (rs) correlations between each pair-wise category of macrohabitat suitability (MS) and soil burn severity (SBS). Correlations are below the diagonal and P-values are above the diagonal; P = 0.5*, P = 0.01**, P = 0.001*** (mod = moderate).
| Category | Watershed ha | MS Low | MS Low-mod | MS Mod | MS Mod-high | MS High | MS Critical | SBS Unburned-very low | SBS Low | SBS Mod | SBS High |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total acres | —— | 0.001*** | 0.001*** | 0.001*** | 0.006** | 0.300 | 0.298 | 0.001*** | 0.001*** | >0.001** | 0.006** |
| Low | 0.650 | —— | 0.031* | 0.731 | 0.268 | 0.185 | 0.332 | 0.001*** | 0.001*** | 0.001** | 0.104 |
| Low-mod | 0.590 | 0.310 | —— | 0.001*** | 0.116 | 0.908 | 0.849 | 0.707 | 0.034* | 0.001** | 0.005** |
| Mod | 0.480 | -0.050 | 0.520 | —— | 0.001*** | 0.135 | 0.466 | 0.222 | 0.019* | 0.002** | 0.084 |
| Mod-high | 0.390 | -0.160 | 0.230 | 0.720 | —— | 0.001*** | 0.024* | 0.032* | 1.015* | 1.102 | 0.716 |
| High | 0.150 | -0.190 | -0.020 | 0.220 | 0.520 | —— | 0.001*** | >0.082 | 0.159 | 0.902 | 0.658 |
| Critical | 0.150 | -0.140 | -0.030 | 0.110 | 0.330 | 0.590 | —— | 0.208 | 0.177 | 0.857 | 0.900 |
| Unburned-very low | 0.630 | 0.470 | 0.060 | 0.180 | 0.310 | 0.250 | 0.190 | —— | 0.001*** | 0.932 | 0.199 |
| Low | 0.850 | 0.600 | 0.310 | 0.340 | 0.350 | 0.210 | 0.200 | 0.790 | —— | 0.001*** | 0.146 |
| Moderate | 0.840 | 0.590 | 0.720 | 0.440 | 0.240 | -0.020 | 0.030 | 0.250 | 0.580 | —— | 0.001*** |
| High | 0.006 | 0.240 | 0.400 | 0.250 | 0.050 | -0.070 | -0.020 | -0.190 | 0.210 | 0.600 | —— |
The SBS map showed that Rusch, Dutch, Lower Big, Canadian, Little, Soldier, East Fork Big French, Rowdy Bar, and Baker creeks, and Drinkwater Gulch had the most ha impacted by the Monument Fire (Table S6). Here there was a significant relationship between total ha in the watershed and total acres of all categories of SBS (Table 4; Fig. 5F). As in categories of habitat suitability, each SBS category also graded into each adjacent SBS class in terms of total ha of habitat affected. Unburned-to-very low burned soil was significantly correlated with Low burned soil, low burned soil was significantly correlated with Moderate burned soil, and Moderate burned soil was significantly correlated with Highly burned soil (Table 4). Ranked correlations between categories of suitable habitat and those of SBS revealed that ha of Low habitat suitability was significantly correlated with Unburned-very low, Low, and Moderately burned areas of SBS. Low-moderate and Moderate habitat suitability’s were significantly correlated with Low, Moderate, and High areas of SBS. Moderate-high suitability was significantly correlated with Unburned-very low and Low burn areas of SBS. However, there was no significant relationship between ha of High or Critical suitability and any category of SBS (Table 3).
Discussion
Impacts of Wildfire and Soil Burn Severity on Habitat Suitability
My study showed that the Monument Fire engulfed a significant portion of the geographic range and suitable macrohabitat of M. setosa at both the landscape- and watershed-levels. This outcome was a function of both total acreage of suitable forest habitat and proportional differences in ha of habitat quality. Total area of forest cover-type vegetation and individual forest stand attributes impacted by the fire was not significantly different from areas that were not burned. Similarly, there were no significant proportional differences between burned and unburned areas in the categories of suitable macrohabitat (i.e., Low–Critical), and only minor differences in the percentages of forest cover-types. There were, however, significant differences in many environmental parameters measured at sample sites between unburned and burned subsets of the original HSM. These differences were a function of the geographic region encompassed by each subset of the original HSM. For example, the bulk of the Monument Fire HSM was located centrally along the northeast boundary of the species range. This region tends to be colder in the winter, and drier and warmer in the summer. By comparison, most of the Non-fire HSM included more southwestern regions of the species range characterized by more moderate temperatures and greater rainfall moving in a coastal westerly direction toward the boundary between Trinity and Humboldt counties.
As expected, the extent to which the fire affected basin watersheds was highly variable. The relationship between total area of the watershed and area of suitable macrohabitat in the Monument Fire HSM was driven by Low to Moderate-high categories of suitability. Low model efficiency, combined with the fact that other variables (i.e., loadings) also influenced the distribution of suitability categories in the 2-dimensional DF plots. Lack of a precise distinction between suitability measures complicated the significance of individual attributes in each set of environmental variables in evaluating the importance and distinctiveness of the suitability criteria modeled. Delineation of watersheds with Moderate-high, High, and Critical macrohabitat suitability provides new knowledge and identification of specific watersheds likely to be most important in post-fire inventory and conservation of the species, particularly as delineated in the analysis of SBS.
I also found significant correlations between levels of SBS and categories of macrohabitat suitability. Overlaying the SBS map onto the Monument Fire HSM revealed that the most extensive areas of soil burn (i.e., Low and Moderate SBS) occurred in areas of Low to Moderate-high habitat suitability. Low SBS generally equates to areas of partial consumption of fine fuels, while litter coverage remains relatively intact on the soil surface. Whereas Moderate SBS typically corresponds to areas in which nearly all soil cover of vegetative litter and fine fuels were consumed or converted to ash (BAER 2022). Conversely, there were few areas of High to Critical habitat suitability affected by high levels of soil burn (i.e., high heat and long burn time). The mosaic burn pattern in High to Critical habitat suitability was a direct function of the small area encompassed by these two “high quality” habitat categories exacerbated by their highly fragmented and widely dispersed geographic distribution. Further, these areas are mostly separated by major topographic and riverine discontinuities in the Greater Trinity Basin; a pattern supported by recent molecular and phylogenetic analyses of M. setosa from throughout its geographic range (Sullivan 2021). Importantly, as with most forest wildfires, it is generally not known to what extent the area and mosaic pattern of soil burn reflected in post-fire SBS maps are a function of regional topography, vegetative characteristics of the landscape, and climate, or the direct anthropogenic actions of fire suppression.
Impacts of Wildfire on Communities of Terrestrial Gastropods
Land snails that occupy layers of forest litter and duff with little or no protection from surface or buried saxicolous habitat are highly vulnerable to fire and changes to their microhabitats (Burke et al. 1999). Bros et al. (2011) found that postfire management produced variation in habitat structure according to different levels of wood removal related to the moist microhabitats provided by accumulation of wood debris on the ground (i.e., trunks, logs, stumps; Appendix IA and IB (PDF); see Sullivan 2022b [Fig. 4 and Appendix 5]). In contrast, taxa that occupied deep subsurface rock-laden fissures or root-wads of large conifer or hardwood trees (i.e., Pacific madrone, tanoak) withstood surface fire to a much greater degree than species that inhabit shallow leaf litter augmented by large woody debris (Burke et al. 1999; Bros et al. 2011; Gaines et al. 2011). For saxicolous species this applies even though high intensity fires may eliminate most individuals in local populations, and their microhabitat requirements and primary food sources. A subsurface rock-dwelling life history strategy is particularly true of M. setosa, in which the most abundant concentrations of individuals generally occupy shady mesic forest conditions that include buried boulder fields and stable slopes (Appendix IC (PDF)). Therefore, understanding the mechanistic causes of these varied responses by terrestrial gastropods is a conservation concern for the management of regional wildlife areas heavily affected by fire (Kiss and Magnin 2002; Santos et al. 2009; Driscoll et al. 2010).
Indeed, excavations of rocky habitat within mesic forest conditions often revealed the presence of live active or estivating snails (and shells) within the spacious and humid catacombs of their subterranean “niche.” Within these moss-covered boulder fields estivating snails and accumulations of empty shells have been found as deep as 1 m underground (see Sullivan 2022b [Fig 6]). Habitation at this depth requires spaces that enabled large-shelled adults to migrate from deep within the recesses of boulder piles and to surface when moisture conditions permitted emergence. Thus, rock matrices may offer Trinity bristle snails protection even from high intensity surface burns, which make specimens less vulnerable to fire and changes to their critical subsurface habitat compared to species that live only amongst forest litter and large woody debris found on the forest floor.
Still, for all forest-dwelling terrestrial gastropods the effect of high levels of soil burn typically destroys (i.e., converts to ash) most vegetative cover, root systems, litter, fine soil fuels, and minerals down to several centimeters deep in the soil (BAER 2022). This process creates bare soil predisposed to the impacts of precipitation and subsequent water runoff and represents the most severe direct and indirect impacts to communities of soil-dwelling terrestrial snails. Modification of water repellency is a much-discussed fire effect because repellency is a natural soil property. Removal of leaf litter, in combination with vegetative cover, frequently results in compaction of soil surface, and alteration and potential destabilization of slope. As evidenced by Driscoll et al. (2021), high intensity fire can increase the severity and the thickness of the water repellent soil which has significant effects to post-fire water runoff in fire-prone areas.
Post-fire Colonization and Dispersal
Endemic populations of M. setosa have a geographic distribution within the Greater Trinity Basin that is highly fragmented topographically, hydrologically, and ecologically (Sullivan 2021 and 2022a). Regional landscape diversity in combination with small population size and low vagility act to restrict geneflow among relic populations. Recent molecular DNA analyses show that these attributes have functioned to facilitate allopatric differentiation leading to elevated levels of taxonomic diversity at the subspecies level (Sullivan 2021). This metapopulation structure of isolated subpopulations may result in altered population dynamics and over the millennium, has undoubtedly caused localized extinction. Colonization of post-fire habitat in mixed conifer forest and riparian ecosystems by relic populations of this species will likely proceed from the distribution of permanent subterrain accretions of large boulders and stable talus in combination with the spatial mosaic of unburned suitable habitat.
For example, densely vegetated areas are expected to create longer burn times, more intense heat, and greater subsurface soil penetration. This process creates a fine-scale mosaic across the landscape in which scattered land snail refugia are present as a result of both the widely distributed mosaic of subsurface rock and as a function of the mosaic pattern of fire behavior, intensity, burn time, and density of vegetation at the soil surface. Kiss and Magnin (2006) and Gongalsky (2011) described similar life-history survival strategies in snails occupying xerophilous boulder ecosystems of the Mediterranean and the Black Sea coast of the Caucasus, which led to lower intensity burns in those areas and higher snail survival. For M. setosa post-fire populations appear highly dependent upon post-fire habitat structure, fire behavior, and the pre-fire distribution of refugial subsurface habitat reflected in the geological and forest ecosystem site histories within the basin.
Suggestions and Recommendations for Post-fire Restoration
Restoration of natural wildfire regimes through fuels reduction and post-fire restoration has emerged as a prime management objective for forests prone to high severity burns within the interior western United States (Huff et al. 2005). Yet such actions are not without their detractors when ostensibly in conflict with ecosystem approaches to conservation of listed, rare, or endemic taxa (Prather et al. 2007; Gaines et al. 2011). Huff et al. 2005 note that land management policies that accelerate fuel reduction using low intensity prescribed burning (e.g., light understory thinning of trees, removal of invasive shrubs; Biswell 1989; Gilliam et al. 2018) are currently being employed with limited scientific information on the ecological consequences for terrestrial animal species throughout the Pacific Northwest (Wright and Agee 2004; Hessburg et al. 2005, 2007). While studies of the effects of wildfire in forest ecosystems on vegetation communities are somewhat abundant for the Klamath Bioregion, research on the impacts of natural fire and prescribed burning on terrestrial gastropods is virtually nonexistent (Agee 2001; Dunk et al. 2004; Severns 2005; Halofsky et al. 2011; Ray and Bergey 2015; Collins et al. 2019; Brand et al. 2020).
Use of GIS-based fire, SBS, and debris-flow maps developed at the scale of drainage basins and stream segments may enable better identification of potential post-fire flashflood debris flows. This methodology is particularly relevant for use in areas of High to Critical suitable habitat (USGS 2022). For M. setosa, the spatial distribution of post-fire populations depends largely on post-fire habitat structure created by scattered refuges as a function of the spatial heterogeneity of the burn and the montage pattern of scattered refuges. Because detection of these subsurface enclaves will prove difficult to locate, increased planning and survey effort via ground-truthing will be required prior to and after prescribed burning, and for post-fire monitoring and mitigation (Hylander 2011). This methodology also applies to communities of terrestrial gastropods when assessing regional or forest-level mollusk biodiversity (Kiss and Magnin 2002). Thus, future post-fire evaluation, inventories, and research into subpopulations will require: 1) documentation of direct fire mortality on individuals and local population abundance; and 2) evaluation of the response of local populations to the effects of opening up the matrix of suitable habitat (i.e., vegetation), which alters fundamentally the levels of light and humidity essential for survival and long-term viability of populations over the next decade.
Across the landscape, fuel treatments that create firesafe conditions for rare or listed terrestrial snails may include a number of specific actions. For endemic populations of M. setosa the most important range-wide conservation actions include: 1) application of low intensity prescribed burning that will create a mosaic of burn conditions (i.e., small and unburned areas < 50 m2), which generally are not hot enough to kill individuals buried underground in their subterranean niches; 2) avoidance of areas of large moss-covered boulders in burning prescriptions that contain or potentially harbor M. setosa and their moist critical microhabitat; and 3) implementation of burns when snails are hibernating, preferrable at or below freezing temperatures.
Conclusions
Understanding how special status species respond to both natural and anthropogenic perturbations in a heterogenous landscape is vital to efficient implementation of future conservation strategies given the expectation that of future climate change will increase the frequency and extent of wildfire (Sanderson et al. 2002; Deguise and Kerr 2006). Understanding the mechanistic causes of this heterogeneous response is a conservation concern for the management of wildfire prone forests and the species that inhabit the Klamath Bioregion (Driscoll et al. 2010; Santos et al. 2009). Here, I present resource and land planners with a landscape-level perspective documenting the extent and potential impact of a major high-severity forest fire on the geographic range and differential extent of burning of suitable for M. setosa. This analysis should prove useful at the watershed level in the development of post-fire surveys, future research, restoration, and conservation strategies given the unique ecology and life history strategy of this endemic species. Results may also serve as a template for evaluating biological diversity in other terrestrial gastropod communities with similar ecologies potentially threatened by wildfire and other anthropogenic habitat degradation within the Klamath Bioregion of northern California.
Acknowledgments
I am grateful to the Unites States Forest Service BAER Program and to Kent Bloom (GIS Specialist) Shasta-Trinity National Forest (Weaverville, CA) for providing the GIS shapefiles of the Monument Fire Soil Burn Severity map. I also thank Dr. Andre DeGeorges and Kent Bloom for providing constructive comments and editorial suggestions on an earlier version of this manuscript. I am particularly appreciative for the financial support provided by the U.S. Fish and Wildlife Service (Project T-21-1 [Project C]) funding portions of this project through the State Wildlife Grant (SWG) Program made available to CDFW.
Literature Cited
- Abele, S. E. 2010. Gastropod diversity in the boreal mixed wood forest of northern Alberta—variation among forest types and response to partial harvesting. Thesis, University of Alberta. Edmonton, Canada.
- Agee, J. K. 1993. Fire Ecology of Pacific Northwest Forests. Island Press, Washington, D.C., USA.
- Agee, J. K. 2001. GOBIG2K: an analysis of fire and mollusk species in California National Forests covered by the Northwest Forest Plan. Final Report, USDA Forest Service, Klamath National Forest. Yreka, CA, USA.
- Airola, D. A. 1988. A guide to the California wildlife habitat relationships system. California Department of Fish and Game, Sacramento, CA, USA. Available from: https://wildlife.ca.gov/Data/CWHR/Wildlife-Habitats/
- Baur, B. 1986. Patterns of dispersion, density, and dispersal in alpine populations of the land snail Arianta arbustorum (L.) (Helicidae). Holarctic Ecology 9(2):117–125.
- Baur, B., and A. Baur. 1990. Experimental evidence for intra- and interspecific competition in two species of rock-dwelling land snails. Journal of Animal Ecology 59:302–315.
- Barrett, J. G. 1966. Climate of Trinity County. United States Department of Agriculture, Soil Conservation Service, Redding, CA, USA.
- Biswell, H. 1989. Prescribed burning in California wildlands vegetation management. University of California Press, Berkeley, CA, USA.
- Brand, M. E., T. W. T. Rechkemmer, S. A. Clark, K. W. McCravy, C. Lydeard, S. T. Meiers, and S. E. Jenkins. 2020. The influence of fire and other environmental factors on terrestrial gastropod species composition in an oak-hickory woodland of west-central Illinois. American Malacological Bulletin 38:39–49.
- Bros, V., G. Moreno-Rueda, and X. Santos. 2011. Does postfire management affect the recovery of Mediterranean communities? The case study of terrestrial gastropods. Forest Ecology and Management 261:611–619.
- Brown, R. T., J. K. Agee, J. F. and Franklin. 2004. Forest restoration and fire: principals in the context of place. Conservation Biology 18:903–912.
- Burke, T. E., J. S. Applegarth, and T. R. Weasma. 1999. Management recommendations for survey and manage terrestrial mollusks. USDA Forest Service, Pacific Northwest
- Region, Portland, OR, USA.
- Burned Area Emergency Response Program (BAER). 2022. USDA Forest Service, Washington, D.C., USA. Available from: https://www.fs.usda.gov/naturalresources/watershed/burnedareas.shtml
- California Department of Fish and Wildlife (CDFW). 1972. California Endangered Species Act (CESA). Available from: https://nrm.dfg.ca.gov/FileHandler.ashx?DocumentID=109406&inline (PDF)
- California Department of Fish and Wildlife (CDFW). 2022. Special Animals. Available from: https://nrm.dfg.ca.gov/FileHandler.ashx?DocumentID=109406&inline (PDF)
- California Natural Diversity Database (CNDDB). 2022. California Department of Fish and Wildlife. Available from: https://wildlife.ca.gov/Data/CNDDB/
- Caturla, R. N., J. Raventós, R. Guàrdia, and R. Vallejo. 2000. Early post-fire regeneration dynamics of Brachypodium retusum Pers. (Beauv.). Acta Oecologica 21:1–12.
- Collins, B. M; S. L. Stephens, and R. A. York. 2019. Perspectives from a long-term study of fuel reduction and forest restoration in the Sierra Nevada. Tree Rings 29:7–9.
- Corder, G. W., and D. I. Foreman. 2014. Nonparametric Statistics: A Step-by-step Approach. John Wiley and Sons, Inc., Hoboken, NJ, USA.
- Cucherat, X., and S. Demuynck. 2008. Sampling strategies and collecting techniques for land and freshwater mollusks. MalaCo 5:244–253.
- Curtis, R. O., and D. D. Marshall. 2000. Why quadratic mean diameter? Western Journal of Applied Forestry 15:137–139.
- Daly, C., R. P. Neilson, and D. L. Phillips. 1994: A statistical-topographic model for mapping climatological precipitation over mountainous terrain. Journal of Applied Meteorology 33:140-158.
- Deguise, I. E., and Keer, J. T. 2006. Protected areas and prospects for endangered species. Conservation in Canada. Conservation Biology 20:48–55.
- Driscoll, D. A, D. B. Lindenmayer, A. F. Bennett, M. Bode, R. A. Bradstock, G. J. Cary, M. F. Clarke, N. Dexter, R. Fensham, G. Friend, M. Gill, S. James, G. Kay, D. A. Keith, C. MacGregor, J. Russel-Smith, D. Salt, J. E. M. Watson, R. J. Williams, and A. York. 2010. Fire management for biodiversity conservation: key research questions and our capacity to answer them. Biological Conservation 143:1928–1939.
- Duncan, N., T. Burke, S. Dowlan, and P. Hohenlohe. 2003. Survey protocol for survey and manage terrestrial mollusk species from the Northwest Forest Plan. USDA Forest Service, Pacific Northwest Region, Portland, OR, USA.
- Duncan, N., 2005. Conservation assessment for Chelan Mountainsnail. USDA Forest Service and USDI Bureau of Land Management, Portland, OR, USA.
- Dunk, J. R. W. J. Zielinski, and H. K. Preisler. 2004. Predicting the occurrence of rare mollusks in Northern California forests. Ecological Applications 14:713–729.
- Fontaine, B., O. Gargominy, and E. Neubert. 2007. Priority sites for conservation of land snails in Gabon: testing the umbrella species concept. Diversity and Distributions 13:725–734.
- Furnish, J., T. Burke T, T. Weasma, J. Applegarth, N. Duncan, R. Monthey, and D. Gowan. 1997. Survey protocol for terrestrial mollusk species from the northwest forest plan, draft version 2.0. USDA Forest Service, Region 6 and the Bureau of Land Management, Oregon and Washington, USA.
- Gaines, W. L., A. L. Lyons, and A. Sprague. 2005. Predicting the occurrence of a rare mollusk in the dry forests of north-central Washington. Northwest Science 79:99–105.
- Gaines, W. L., A. L. Lyonsam K. Weaver, and A. Spraguea. 2011. Monitoring the short-term effects of prescribed fire on an endemic mollusk in the dry forests of the eastern Cascades, Washington, USA. Forest Ecology and Management 261:1460–1465.
- Garrison, B. A., M. D. Parisi, K. W. Hunting, T. A. Giles, J. T. McNerney, R. G. Burg, K. J. Sernka, S. L. Hooper. 2002. Training manual for California Wildlife Habitat Relationships System CWHR database version 8.0. 9th edition. California Wildlife Habitat Relationships Program, California Department of Fish and Game, Sacramento, CA, USA.
- Gilliam, M. E., W. T. Rechkemmer, K. W. McCravy, and S. E. Jenkins. 2018. The influence of prescribed fire, habitat, and weather on Amblyomma americanum (Ixodida: Ixodidae) in West-Central Illinois, USA. Insects 9:36.
- Gongalsky, K. 2011. The spatial distribution of large soil invertebrates on burned areas in xerophilous ecosystems of the Black Sea coast of the Caucasus. Arid Ecosystems 1:260–266.
- Goodchild, M., F. W. Davis, M. Painho, and D. M. Storns. 1991. The use of vegetation maps and geographic information systems for assessing conifer lands in California. Technical Report 91-23, National Center for Geographic Information and Analysis, University of California, Santa Barbara, CA, USA.
- Gotmark, F., T. Von Proschwitz, and N. Franc. 2008. Are small sedentary species affected by habitat fragmentation? Local vs. landscape factors predicting species richness and composition of land mollusks in Swedish conservation forests. Journal of Biogeography 35:1062–76.
- Halofsky, J. E., D. C. Donato, D. E. Hibbs, J. L. Campbell, M. Donaghy Cannon, J. B. Fontaine, J. R. Thompson, R. G. Anthony, B. T. Bormann, L. J. Kayes, B. E. Law, D. L. Peterson, and T. A. Spies. 2011. Mixed-severity fire regimes: lessons and hypotheses from the Klamath-Siskiyou Ecoregion. Ecosphere 2:1–19.
- Hansson, L. 1991. Dispersal in the edible snail, Helix pomatia: a test case for present generalizations. Acta Oecologica 12:761–769.
- Helms, J. 1998. The Dictionary of Forestry. Society of American Foresters, Washington, D.C., USA.
- Hessburg, P. E., B. G. Smith, and R, B, Slater. 1999. Detecting change in forest spatial pattern from reference conditions. Ecological Applications 9:1232–1252.
- Hessburg, P. F., J. K. Agee, and J. F. Franklin. 2005. Dry forests and wildland fires of the inland Northwest, USA: contrasting the landscape ecology of the pre-settlement and modern eras. Forest Ecology and Management 211:117–139.
- Hessburg, P. F., K. M. James, and R. B. Salter. 2007. Re-examining fire severity relations in pre-management era mixed conifer forests: inferences from landscape patterns of forest structure. Landscape Ecology 22:5–24.
- Huff, M. H, N. E. Seavy, J. D. Alexander, and C. J. Ralph. 2005. Fire and birds in maritime Pacific northwest. Studies in Avian Biology 30:46–62.
- Hylander, K. 2011. The response of land snail assemblages below aspens to forest fire and clear-cutting in Fennoscandian boreal forests. Forest Ecology and Management 261:1811–1819.
- Kiss, L. M., and F. M. Magnin. 2002. The impact of fire on land snail communities in the French Mediterranean region: preliminary results. Pages 197–213 in L. Trabaud and R. Prodon, editors. Fire and Biological Processes. Backhuys Publishers, Leiden, Netherlands.
- Kiss, L., and F. Magnin. 2003. The impact of fire on some Mediterranean land snail communities and patterns of post-fire recolonization. Journal of Molluscan Studies 69:43–53.
- Kiss, L., F. Magnin, and F. Torre. 2004. The role of landscape history and persistent biogeographical patterns in shaping the responses of Mediterranean land snail communities to recent fire disturbances. Journal of Biogeography 31:145–157.
- Kiss, L., and F. Magnin. 2006. High resilience of Mediterranean land snail communities to wildfires. Biodiversity and Conservation 15:2925–2944.
- Martin, K., and M. Sommer. 2004. Relationships between land snail assemblage patterns and soil properties in temperate-humid forests. Journal of Biogeography 31:531–545.
- Mayer K. E, and W. F. Laudenslayer. 1988. A guide to the wildlife habitats of California. California Department of Forestry and Fire Protection, Sacramento, CA, USA.
- Moretti, M., F. de Bello, S. P. M. Roberts. and S. G. Potts. 2009. Taxonomical vs. functional responses of bee communities to fire in two contrasting climatic regions. Journal of Animal Ecology 78:98–108.
- Nekola, J. C. 2002. Effects of fire management on the richness and abundance of central North American grassland land snail faunas. Animal Biodiversity and Conservation 25.2:53–66.
- Nelder, J. A., and R. W. M. Wedderburn. 1972. Generalized Linear Models. Journal of the Royal Statistical Society, Series A 135:370–384.
- Noss, R. F., J. F. Franklin, W. L. Baker, T. Schoennagel, and P. B. Moyle. 2006. Managing fire-prone forests in the western United States. Frontiers in Ecology and the Environment 4:481–487.
- Parker, I., and W. J. Matyas. 1981. CALVEG: a classification of California vegetation. USDA Forest Service, Regional Ecology Group, San Francisco, CA, USA.
- Pausas, J. G., and M. Verdú. 2005. Plant persistence traits in fire-prone ecosystems of the Mediterranean Basin: a phylogenetic approach. Oikos 109:196–202.
- Prather, J. W., R. F. Noss, and T. D. Sisk. 2008. Real versus perceived conflicts between restoration of ponderosa pine forests and conservation of the Mexican spotted owl. Forest Policy and Economics 10:140–150.
- Raheem, D. C., F. Naggs, R. C. Preece, Y. Mapatuna, L. Kariyawasam, and P. Eggleton. 2008. Structure and conservation of Sri Lankan land-snail assemblages in fragmented lowland rainforest and village home gardens. Journal of Applied Ecology 45:1019–1028.
- Ray, E. J., and E. A. Bergey. 2015. After the burn: Factors affecting land snail survival in post-prescribed-burn woodlands. Journal of Molluscan Studies 81:44–50.
- Razali, N., and Y. B. Wah. 2011. Power comparisons of Shapiro-Wilk, Kolmogorov-Smirnov, Lilliefors and Anderson-Darling tests. Journal of Statistical Modeling and Analytics 2:21–33.
- Rosner, B. R. J. Glynn, and M. T. Lee. 2006. The Wilcoxon signed rank test for paired comparisons of clustered data. Biometrics 62:185–92.
- Roth, B. 1978. Biology and distribution of Monadenia setosa Talmadge. Report to U.S. Forest Service, Shasta-Trinity National Forest, Redding, CA, USA.
- Roth, B., and L. L. Eng. 1980. Distribution, ecology, and reproductive anatomy of a rare land snail. Monadenia setosa, Talmage. California Fish and Game 66:4–16.
- Roth, B. 1982. Life history studies and distribution of Monadenia setosa. Report submitted to USDA Forest Service, Shasta-Trinity National Forest, Redding CA, USA.
- Roth, B., and P. H. Pressley. 1986. Observations on the range and natural history of Monadenia setosa (Gastropoda: Pulmonate) in the Klamath Mountains, California, and the taxonomy of some related species. The Veliger 29:169–182.
- Sanderson, E. W., J. Malanding, M. A. Levy, K. H. Redford, A. V. Wannebo, and G. Woolmer. 2002. The human footprint and the last of the wild. BioScience 52:891–904.
- Santos, X., V. Bros, and A. Miño. 2009. Recolonization of a burned Mediterranean area by terrestrial gastropods. Biodiversity Conservation 18:153–3165.
- Sawyer, J. O., and T. Keeler-Wolfe. 1995. A Manual of California Vegetation. California Native Plant Society, Sacramento, CA, USA.
- Severns, P. M. 2005. Response of a terrestrial mollusk community to an autumn prescribed burn in a rare wetland prairie of western Oregon, USA. Journal of Molluscan Studies 71:181–187.
- Sullivan, R. M. 2021. Phylogenetic relationships among subclades within the Trinity bristle snail species complex, riverine barriers, and re-classification. California Fish and Wildlife Journal, CESA Special Issue:107–145.
- Sullivan, R. M. 2022a. Macrohabitat suitability models for the Trinity bristle snail (Monadenia setosa) in the Greater Trinity Basin of Northern California. California Fish and Wildlife Journal 108:e2. https://doi.org/10.51492/cfwj.108.2
- Sullivan, R. M. 2022b. Microhabitat characteristics and management of the Trinity bristle snail in the Greater Trinity Basin of northern California. California Fish and Wildlife Journal 108:e3. https://doi.org/10.51492/cfwj.108.3
- Talmadge, R. R. 1952. A bristled Monadenia from California. The Nautilus 66:47–50.
- United States Geological Survey (USGS). 2022. Emergency assessment of post-fire debris-flow hazards. Available from: https://landslides.usgs.gov/hazards/postfire_debrisflow/
- U.S. Forest Service (USFS). 1981. CALVEG: A Classification of California Vegetation. Pacific Southwest Region, Regional Ecology Group, San Francisco CA, USA.
- Walton, M. L. 1963. Length of life in west American land snails. The Nautilus 76:127–131.
- Welsh, H. H. 1994. Bioregions: An ecological and evolutionary perspective and a proposal for California. California Fish and Game 80:97–124.
- Whelan, R. J. 1995. The Ecology of Fire. Cambridge University Press, Cambridge, UK.
- Wright, C. S., and J. L. Agee. 2004. Fire and vegetation history in the eastern Cascade mountains, Washington. Ecological Applications 14:443–459.