

FULL RESEARCH ARTICLE
Brian Cypher* and Erica Kelly
California State University, Stanislaus, One University Circle, Turlock, CA 95382, USA![]() https://orcid.org/0000-0002-7349-545X (BC)
https://orcid.org/0000-0002-7349-545X (BC)![]() https://orcid.org/0000-0002-3033-1607 (EK)
https://orcid.org/0000-0002-3033-1607 (EK)
*Corresponding Author: bcypher@esrp.csustan.edu
Published 28 August 2025 • doi.org/10.51492/cfwj.111.14
Abstract
The immense ecological plasticity of coyotes (Canis latrans) facilitates their use of anthropogenically altered habitats including urban areas. We analyzed scat samples to compare food item use by coyotes in urban, peri-urban, and non-urban zones in and around the city of Bakersfield, California. Coyotes consumed primarily rabbits and rodents in all three zones although species composition varied among the zones. Other important foods included birds and invertebrates in the urban zone, commercial grapes in the peri-urban zone, and birds and fruits in the non-urban zone. Frequency of occurrence of anthropogenic items was not statistically different between zones but exhibited an increasing trend from the non-urban zone to the urban zone. However, coyotes in the urban zone do not appear to be dependent on these items. The relatively low use of anthropogenic foods may reduce the potential for human-coyote conflicts in Bakersfield.
Key words: Canis latrans, coyotes, food habits, human-wildlife conflict, San Joaquin Valley, urban environment
| Citation: Cypher, B., and E. Kelly. 2025. Coyote food item use along an urban gradient in the southern San Joaquin Valley, California. California Fish and Wildlife Journal 111:e14. |
| Editor: Matt Meshiry, Wildlife Branch |
| Submitted: 15 May 2025; Accepted: 1 July 2025 |
| Copyright: ©2025, Cypher and Kelly. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Funding: This project was funded by the CSU-Stanislaus Endangered Species Recovery Program. |
| Competing Interests: The authors have not declared any competing interests. |
Introduction
Coyotes (Canis latrans) exhibit tremendous adaptability that is a consequence of their immense ecological plasticity (Bekoff and Gese 2003). This has helped this species extensively expand its range (Laliberte and Ripple 2004; Hody and Kays 2018) and readily occupy anthropogenically altered habitats including urbanized environments (MacCracken 1982; Gehrt and Riley 2010). An aspect of this plasticity is an ability to exploit a wide variety of foods (Jensen et al. 2022). In urban areas, this can include anthropogenic items (Fedriani et al. 2001; Newsome et al. 2015) and in some locations coyotes may develop a dependency on such items potentially leading to human-coyote conflict situations (Gehrt and Riley 2010; Lukasik and Alexander 2012; Murray et al. 2015).
In this study, we examined food item use by coyotes in the city of Bakersfield as well as nearby peri-urban and non-urban sites. Our objectives were to determine (1) whether coyote food item use differed among urbanization zones and (2) the extent to which coyotes were using anthropogenic items, particularly in the urban zone.
Methods
Study Area
We conducted this study in the southern San Joaquin Valley in central California in and around the city of Bakersfield. Bakersfield encompasses approximately 390 km2 and has a human population exceeding 403,000 as of the 2020 census (https://en.wikipedia.org/wiki/Bakersfield,-_California). We collected coyote scats at four sites in three zones of urbanization: urban, peri-urban, and non-urban (Fig. 1). The two urban sites were the campus of the California State University-Bakersfield (CSUB; 35.349, –119.103) and the Bakersfield Renewable Fuels complex (BRF; 35.371, –119.079). The peri-urban site was a patchwork of open and commercial lands on the north edge of Bakersfield by the Meadows Field Airport (MFA; 35.444, –119.047). The non-urban site was Kern Water Bank conservation lands located approximately 8 km west of the western edge of Bakersfield (KWB; 35.326, –119.233).

CSUB is surrounded by urban land uses consisting primarily of commercial and residential developments. On the campus, irrigated lawns and landscaping are present around buildings and on athletic fields. However, large portions of the campus are unirrigated and covered by a dense growth of ruderal plants, particularly non-native species such as red brome (Bromus madritensis), wild barley (Hordeum murinum), black mustard (Brassica nigra), and puncture vine (Tribulus terrestris). BRF is an industrial complex with large open spaces that are periodically scraped to clear vegetation. Small patches of ruderal vegetation are present consisting primarily of red brome and wild barley. Some irrigated lawns and landscaping are present around buildings in the MFA area. Ground cover in open spaces varies from sparse to dense and consists primarily of red brome, wild barley, black mustard, and wild oats (Avena spp.). Widely scattered shrubs consist primarily of desert saltbush (Atriplex polycarpa) and bladderpod (Cleome isomeris). The vegetation community on the KWB site is largely saltbush scrub that is generally consistent with the Allscale Series (Sawyer and Keeler-Wolf 1995). Ground cover is sparse to dense and consists of red brome, wild barley, and vinegarweed (Trichostema lanceolatum). Scattered shrubs are present and consist of desert saltbush, goldenbush (Isocoma spp.), and honey mesquite (Prosopis glandulosa). A few water recharge ponds are present. Sheep grazing occurs on the site in the spring and cattle are present in some years as well.
The regional climate is Mediterranean, characterized by hot, dry summers and cool, wet winters. Mean maximum and minimum temperatures were 35°C and 18°C, respectively, in summer, and 17°C and 5°C, respectively, in winter. Annual precipitation averages 15 cm and occurs primarily as rain falling between October and April (NOAA 2020).
Scat Collection and Analysis
We collected coyote scats from August 2021 to July 2022 opportunistically in conjunction with other field activities. Collections were conducted on two occasions in the urban zone, three occasions in the peri-urban zone, and two occasions in the non-urban zone. Our goal was to collect a minimum of 50 scats from each zone. We located scats by searching along unpaved roads and in open spaces with little or no vegetation. Bobcats (Lynx rufus) were present in low numbers in the peri-urban and non-urban zones, and scats suspected of being from bobcats (e.g., highly segmented with blunt ends; R. Roberts, CDFW Bobcat Project Coordinator, pers. comm.) were avoided. Domestic dogs (Canis lupus familiaris) were present in the peri-urban zone and scats suspected of being from dogs (e.g., obvious dog food material, no natural items) were also avoided. We limited collections to scats considered to be fresh (e.g., no bleaching).
We placed individual scats in paper bags labeled with the date and location coordinates. Scats were oven-dried at 60℃ for ≥24 hr to neutralize any zoonotic parasite eggs and cysts. We then placed the scats in individual nylon bags, washed in a washing machine to remove soluble materials, and dried in a tumble dryer. The remaining undigested material was examined to identify food items. Mammalian remains (e.g., hair, teeth, bones) were identified using macroscopic (e.g., length, texture, color, banding patterns) and microscopic (e.g., cuticular scale patterns) characteristics of hairs (Moore et al. 1974) and by comparing teeth and bones to reference guides (Glass 1981; Roest 1986) and specimens. Other vertebrates were identified to class and invertebrates to order, based on feathers, scales, and exoskeleton characteristics and comparison to reference specimens. Fruits were identified at least to genus based on seed characteristics (Young and Young 1992).
We combined data from the two sites in the urban zone for analyses. Frequency of occurrence (FOO) of each item (number of scats with the item divided by the total number of scats) was determined for each zone. Most scats contained multiple items, and therefore, the sum of the individual FOOs for a given zone will be greater than 100%. For statistical analyses, we grouped items into seven categories: rabbit, rodent, bird, reptile, invertebrate, fruit, and anthropogenic. We considered any items present in the environment due to human activities to be “anthropogenic” items, including domestic animals, crops, ornamental fruits, and trash. We conducted contingency table analyses and a chi-square test for independence to compare FOO among zones for each food category. Yate’s correction-for-continuity was applied in all 2×2 contingency tables. A Fisher’s Exact Test was employed for any 2×2 tables that included a cell with less than 5 occurrences (Gotelli and Ellison 2004). We adopted a more lenient alpha level of 0.1 to better identify trends that warranted further investigation (Gotelli and Ellison 2004). Shannon diversity indices (H¢) were calculated for each zone using the equation:
H’ = (N log N – ∑ni log ni)/N
where N is the total number of occurrences of all items and ni is the number of occurrences of item i (Brower and Zar 1984).
Results
We collected 104 coyote scats in the urban zone, 59 in the peri-urban zone, and 68 in the non-urban zone. Items positively identified in scats included rabbit (black-tailed jackrabbit [Lepus californicus] or desert cottontail [Sylvilagus audubonii]), Heermann’s kangaroo rat (Dipodomys heermanni), San Joaquin pocket mouse (Perognathus inornatus), Gambel’s deer mouse (Peromyscus gambelii), house mouse (Mus musculus), rat (Rattus spp.), California ground squirrel (Otospermophilus beecheyi), pocket gopher (Thomomys bottae), domestic sheep (Ovis aries), domestic cat (Felis silvestris catus), unidentified bird and eggshells (Class Aves), unidentified snake (Order Squamata), darkling beetle (Eleodes spp.), June beetle (Phyllophaga spp.), unidentified beetle larvae (Order Coleoptera), earwig (Forficula auricularia), botfly (Family Oestridae), scorpion (Order Scorpiones), solpugid (Order Solifugae), snail (Helix spp.), grape (Vitis spp.), honey mesquite, palm (Family Palmaceae), pistachio (Pistacia vera), walnut (Juglans regia), olive (Olea spp.), and miscellaneous items (e.g., foil, paper towel, food wrapper, plastic pieces, piece of rope, leather glove, wood pieces). Individual items occurring frequently in scats from each of the zones (Table 1) included: rabbits, ground squirrels, gophers, birds, and darkling beetles in the urban zone; rabbits, ground squirrels, and grapes in the peri-urban zone; and rabbits, kangaroo rats, deer mice, gophers, birds, and honey mesquite in the non-urban zone.
Table 1. Frequency of occurrence (%) by urbanization zone (urban, peri-urban, and non-urban) of food items in coyote scats from urban, peri-urban, and non-urban zones in the southern San Joaquin Valley during August 2021–July 2022.
| Items | Urban | Peri-urban | Non-urban |
| Rabbit | 23.1 | 62.7 | 39.7 |
| Kangaroo rat | – | – | 19.1 |
| Pocket mouse | 2.9 | 5.1 | 4.4 |
| Deer mouse | 1.0 | – | 14.7 |
| House mouse | – | 1.7 | – |
| Rat | 1.0 | – | – |
| Ground squirrel | 51.9 | 16.9 | 1.5 |
| Pocket gopher | 16.3 | 8.5 | 20.6 |
| Domestic sheep | – | – | 1.5 |
| Domestic cat | 7.7 | – | 1.5 |
| Bird | 14.4 | 3.4 | 11.8 |
| Snake | 3.8 | 1.7 | 2.9 |
| Darkling beetle | 13.5 | 5.1 | 2.9 |
| June beetle | 1.0 | – | – |
| Beetle larva | – | 1.7 | – |
| Earwig | 1.0 | – | 1.5 |
| Bot fly | – | – | 1.5 |
| Scorpion | 1.9 | – | – |
| Solpugid | 1.9 | – | – |
| Snail | – | 1.7 | – |
| Grape | – | 10.2 | – |
| Honey mesquite | – | – | 25.0 |
| Ornamental palm | – | 1.7 | – |
| Pistachio | – | – | 7.4 |
| Walnut | 1.0 | – | – |
| Olive | 1.0 | – | – |
| Trash | 5.8 | 1.7 | – |
By category, the most frequently occurring items in scats were: rodents, invertebrates, and rabbits in the urban zone; rabbits and rodents in the peri-urban zone; and rodents, rabbits, and fruit in the non-urban zone (Fig. 2). Frequency of occurrence of rabbit, rodent, invertebrate, bird, and fruit differed among zones (Table 2). FOO for rabbit was highest in the peri-urban zone and lowest in the urban zone, FOO for rodent was highest in the urban zone and lowest in the peri-urban zone, FOO for invertebrate was highest in the urban zone and similar between the peri-urban and non-urban zones, FOO for bird was highest in the urban and non-urban zones and lowest in the peri-urban zone, and FOO for fruit was highest in the non-urban zone and lowest in the urban zone (Fig. 2). Domestic sheep, domestic cat, grape, palm, pistachio, walnut, olive, and the miscellaneous items all were considered to be anthropogenic items. Although not statistically different among zones, the FOO of anthropogenic items was highest in the urban zone and lowest in the non-urban zone. The diversity of items consumed by coyotes was similar among the urbanization zones, based on the categories (Table 2).
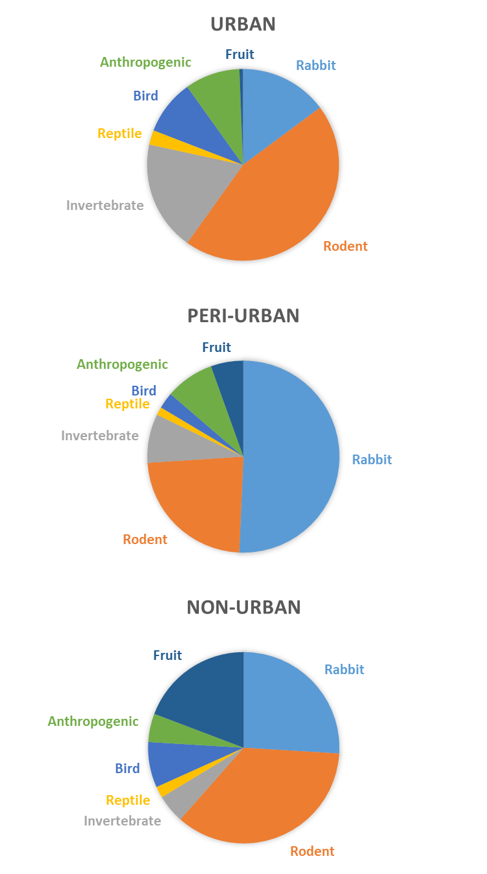
Table 2. Frequency of occurrence (%) by urbanization zone (urban, peri-urban, and non-urban) of food items by category and diversity index (H’) in coyote scats from urban, peri-urban, and non-urban zones in the southern San Joaquin Valley during August 2021–July 2022. Values within a category with the same capital letter did not differ statistically (P > 0.1).
| Categories | Urban | Peri-urban | Non-urban | χ22 | P |
|---|---|---|---|---|---|
| Rabbit | 23.1 C | 62.7 A | 39.7 B | 25.18 | < 0.01 |
| Rodent | 70.2 A | 28.8 C | 54.4 B | 26.05 | < 0.01 |
| Bird | 14.4 A | 3.4 B | 11.8 AB | 4.84 | 0.09 |
| Reptile | 3.8 A | 1.7 A | 2.9 A | 0.60 | 0.74 |
| Invertebrate | 28.8 A | 10.2 B | 7.4 B | 16.13 | < 0.01 |
| Fruit | 1.0 C | 6.8 B | 29.4 A | 35.82 | < 0.01 |
| Anthropogenic | 14.4 A | 10.2 A | 7.4 A | 2.15 | 0.34 |
| H’ | 0.66 | 0.61 | 0.69 | — | — |
Discussion
Our data set is limited in that we were only able to collect scats for one year. Item availability can vary markedly in the non-urban zone due to annual precipitation levels. However, we feel that the data from the urban zone are adequately representative of food item use by coyotes, particularly with regards to proportional use of anthropogenic items relative to natural items. Conditions such as moisture availability tend to be relatively stable in the urban zone due to factors such as year-round landscape irrigation.
Dietary diversity based on food item categories was similar among the three urbanization zones. Likewise, Fedriani et al. (2001) reported that dietary diversity was similar for three areas with different residential development (24%, 10%, and 2%) in southern California. In our study and the Fedriani et al. (2001) study, coyotes primarily consumed rabbits and rodents in all three urbanization zones. Rodents and rabbits commonly are identified as primary foods of urban coyotes (Gehrt and Riley 2010; Duncan et al. 2020). However, in our study the particular species of rabbits and rodents consumed varied among the zones. Based on observations, black-tailed jackrabbits were the predominant species in the non-urban zone and likely comprised the majority of rabbit occurrences in scats from this zone. This is consistent with Ferrel et al. (1953) who found that most of the rabbits consumed by coyotes in the Central Valley of California were black-tailed jackrabbits. However, this species was rare in the urban zone where desert cottontails were primarily observed. Both black-tailed jackrabbits and desert cottontails were commonly observed in the peri-urban zone, and this might explain the significantly higher FOO for rabbit in scats from the peri-urban zone.
Similar to rabbits, rodents were important food item in all zones, but the species consumed differed among zones. Gophers were consumed in all zones. Based on sign (e.g., mounds), gophers were abundant in all zones. California ground squirrels were rarely consumed in the non-urban zone and were more commonly consumed in the peri-urban zone. However, they were frequently consumed in the urban zone and occurred in over half of the scat samples. Ground squirrels are extremely abundant throughout Bakersfield, probably as a result of the extensive irrigation of lawns, landscaping, golf courses, parks, school campuses, and other areas. In the non-urban zone, coyotes commonly consumed kangaroo rats and deer mice, both of which are abundant in the arid habitat of this zone and are common food items for coyotes in the San Joaquin Valley (Cypher et al. 1994; Spiegel et al. 1996; Cypher et al. 2009).
Of note, the occurrence of synanthropic commensal rodents (i.e., rat, house mouse) in coyote scats was rare with one occurrence each in the urban and peri-urban zones. Consumption of commensal rodents was posited to be the likely source of exposure to rodenticides for coyotes in urbanized areas in southern California (Stapp et al. 2025). In that study, 98.1% of 365 urban coyotes sampled had detectable concentrations of first and second generation anticoagulant rodenticides. In our study, coyotes in the urban zone commonly consumed California ground squirrels and gophers, both of which are frequent targets of pest control efforts (CSUS Endangered Species Recovery Program, unpublished data). The exposure rates of coyotes to rodenticides in Bakersfield is not known.
Eight occurrences (7.7% FOO) of domestic cat were recorded in coyote scats from the urban zone. Whether the cats consumed were companion or feral animals and whether they were consumed as live prey or carrion (e.g., road killed) are unknown. Gehrt and Riley (2010) reported that the occurrence of domestic cat in the diet of urban coyotes commonly is low (<5%) although two exceptions were results from King County, Washington (12.6%; Quinn 1997) and San Diego and Orange County, California (15.5%; Larson et al. 2015). Coyotes killing cats is a significant human-coyote conflict issue in some locations leading to reduced tolerance of coyotes (Gehrt and Riley 2010) and even the initiation of control measures (Baker and Timm 2017). The one occurrence of cat in a scat from the non-urban zone likely was a result of predation on a feral cat as no human residences were present in this zone.
Domestic sheep in one scat from the non-urban zone could have been a result of predation or scavenging. Sheep are grazed in that zone each spring and lambs would be easy prey for coyotes, although most herds are guarded by a shepherd and multiple herding dogs. Also, any sheep that die in the field are usually left there resulting in scavenging opportunities. Birds in scats could not be identified as mostly small, gray downy feathers were found. However, the species consumed by coyotes likely differed among zones, particularly the urban and non-urban zones where the FOO for birds was highest. Similarly, we were not able to identify the particular snake species consumed from the scales that were present in scats.
Interestingly, coyotes more frequently consumed invertebrates, particularly beetles, in the urban zone compared to the other zones. It is unknown whether this was a function of relative availability, either among zones or relative to the availability of other food items. Coyotes readily consume fruits when they are available. Fruits consumed by coyotes in the urban zone likely were from landscape plantings or food gardens. A number of commercial table grape vineyards were present near the peri-urban zone and these likely were the source of the grape seeds found in the scats. Honey mesquite was dispersed throughout the non-urban zone and coyotes commonly consume the seed pods in the spring when the pods are developing and still have some fleshy endocarp prior to maturing and drying.
In our review of other studies, the FOO of anthropogenic food items in urban coyote scats varied from 0% (Thompson 2014) to 72% (Caspi et al. 2025). However, quantifying use of anthropogenic items by urban coyotes and comparing results among studies is challenging. As has been abundantly demonstrated through isotopic and molecular analyses (e.g., Newsome et al. 2010, 2015; Caspi et al. 2025), the FOO of anthropogenic food items in scats as determined by macro-analysis is likely an underestimate of actual use because many of these items (e.g., processed snack foods) have few, if any, recognizable indigestible components. Also, the items categorized as anthropogenic vary among studies. We included all items associated with the presence of humans including crops, ornamental fruits, domestic animals, and trash (e.g., food wrappers). In other studies, various combinations of the above are included, and in some studies only items associated with human food items (e.g., food wrappers) are included. Thus, the definition of anthropogenic items needs to be carefully considered when comparing results among studies.
In our study, the FOO of anthropogenic items exhibited an increasing, although not statistically significant, trend from non-urban zone to urban zone. This trend is not surprising given that such items are likely to be more abundant in the urban zone where human activity is higher. A similar trend was reported in southern California where the seasonal FOO of anthropogenic items in coyote scats varied from trace to 3%, 4 to 6%, and 14 to 25% in areas with low, medium, and high residential development, respectively (Fedriani et al. 2001). In our study, coyotes in the urban zone did not appear to be extensively using or reliant upon anthropogenic food items. Just six of the 104 scats (6%) from the urban zone had only anthropogenic items: four of these had cat remains and two had food wrapper remnants. This is a positive result because reliance on anthropogenic foods can lead to habituation and increased human-coyote conflicts (White and Gehrt 2009; Lukasik and Alexander 2011) including attacks on people (Carbyn 1989; Gehrt and Riley 2010; Baker and Timm 2017).
We documented variation in coyote foraging patterns in response to relative item availability mediated by landscape urbanization. This variation elucidates adaptive responses that facilitate coyote persistence in anthropogenically altered landscapes. Based on our results, foraging patterns of coyotes inhabiting Bakersfield currently have low potential for human-coyote conflicts. Outreach programs emphasizing the importance of not provisioning coyotes with anthropogenic foods could help minimize future conflicts.
Acknowledgments
We thank T. Westall, N. Deatherage, A. Gabaldon, and students from the Hall Wildlife Lab at the California State University, Bakersfield for assistance with scat collection. This project was funded by the CSU-Stanislaus Endangered Species Recovery Program. We also thank Bakersfield Renewable Energy, Kern Water Bank Authority, California State University-Bakersfield, and Meadows Field for providing access to their properties to collect scat samples. We thank two anonymous reviewers for providing comments that helped to improve our manuscript.
Literature Cited
- Baker, R. O., and R. M. Timm. 2017 Coyote attacks on humans, 1970–2015: implications for reducing the risks. Human-Wildlife Interactions 2(2):art3.
- Bekoff, M., and E. M. Gese. 2003. Coyote (Canis latrans). Pages 467–481 in G. A. Feldhamer, B. C. Thompson, and J. A. Chapman, editors. Wild Mammals of North America: Biology, Management, and Conservation. 2nd Edition. John Hopkins University Press, Baltimore, MD, USA.
- Brower, J. E., and J. H. Zar. 1984. Field and laboratory methods for general ecology. Wm. C. Brown Publishers, Dubuque, IA, USA.
- Carbyn, L. N. 1989. Coyote attacks on children in western North America. Wildlife Society Bulletin 17:444–446.
- Caspi, T., M. G. Serrano, S. L. Vanderzwan, J. Kessler, C. J. Schell, and B. N. Sacks. 2025. Impervious surface cover and number of restaurants shape diet variation in an urban carnivore. Ecosphere 16:e70152.
- Cypher, B. L., C. D. Bjurlin, and J. L. Nelson. 2009. Effects of roads on endangered San Joaquin kit foxes. Journal of Wildlife Management 73:885–893.
- Cypher, B. L., K. A. Spencer, and J. H. Scrivner. 1994. Food item use by coyotes at the Naval Petroleum Reserves, California. Southwestern Naturalist 39:91–95.
- Duncan, N., O. Asher, M. Weckel, C. Nagy, C. Henger, F. Yau, and L. Gormanzano. 2020. Baseline diet of an urban carnivore on an expanding range front. Journal of Urban Ecology 6(1):juaa021.
- Fedriani, J. M., T. K. Fuller, and R. M. Sauvajot. 2001. Does availability of anthropogenic food enhance densities of omnivorous mammals? An example with coyotes in southern California. Ecography24: 325–331.
- Ferrel, C. M., H. R. Leach, and D. F. Tillotson. 1953. Food habits of the coyote in California. California Fish and Game 39:301–341.
- Gehrt, S. D., and S. P. D. Riley. 2010. Coyotes (Canis latrans). Pages 78–95 in S. D. Gehrt, S. P. D. Riley, and B. L. Cypher, editors. Urban Carnivores: Ecology, Conflict, and Conservation. John Hopkins University Press, Baltimore, MD, USA.
- Glass, B. P. 1981. Key to the Skulls of North American Mammals. Oklahoma State University, Stillwater, OK, USA.
- Gotelli, N. J. and A. M. Ellison. 2004. A Primer of Ecological Statistics. Sinauer Associates, Inc., Sunderland, MA, USA.
- Hody, J. W., and R. Kays. 2018. Mapping the expansion of coyotes (Canis latrans) across North and Central America. ZooKeys759: 81–97.
- Jensen, A. J., C. J. Marneweck, J. C. Kilgo, and D. S. Jachowski. 2022. Coyote diet in North America: geographic and ecological patterns during range expansion. Mammal Review 52:480–496.
- Laliberte, A. S., and W. J. Ripple. 2004. Range contractions of North American carnivores and ungulates. Bioscience54:123–138.
- Larson, R. N., D. J. Morin, I. A. Wierzbowska, and K. R. Crooks. 2015. Food habits of coyotes, gray foxes, and bobcats in a coastal southern California urban landscape. Western North American Naturalist 75:339–347.
- Lukasik, V. M., and S. M. Alexander. 2011. Human-coyote interactions in Calgary, Alberta. Human Dimensions of Wildlife 16:114–127.
- Lukasik, V. M., and S. M. Alexander. 2012. Spatial and temporal variation of coyote (Canis latrans) diet in Calgary, Alberta. Cities and the Environment 4(1):art8.
- MacCracken, J. G. 1982. Coyote food in a southern California suburb. Wildlife Society Bulletin 10:280–281.
- Moore, T. D., L. E. Spence, and C. E. Dugnolle. 1974. Identification of the dorsal hairs of some animals of Wyoming. Wyoming Game and Fish Department, Cheyenne, WY, USA.
- Murray, M., A. Cembrowski, A. D. M. Latham, V. M. Lukasik, S. Pruss, and C. C. St. Clair. 2015. Greater consumption of protein-poor anthropogenic food by urban relative to rural coyotes increases diet breadth and potential for human–wildlife conflict. Ecography 38:1235–1242.
- National Oceanic and Atmospheric Administration (NOAA). 2020. Local climatological data, Bakersfield, California. National Climatological Data Center, Ashville, NC, USA.
- Newsome, S. D., H. M. Garbe, E. C. Wilson, and S. D. Gehrt. 2015. Individual variation in anthropogenic resource use in an urban carnivore. Oecologia 178:115–28.
- Newsome, S. D., K. Ralls, C. Van Horn Job, M. L. Fogel, and B. L. Cypher. 2010. Stable isotopes evaluate exploitation of anthropogenic foods by the endangered San Joaquin kit fox (Vulpes macrotis mutica). Journal of Mammalogy 91:1313–1321.
- Quinn, T. 1997. Coyote (Canis latrans) food habits in three urban habitat types of western Washington. Northwest Science 71:1–5.
- Roest, A. I. 1986. A Key-Guide to Mammal Skulls and Lower Jaws. Mad River Press, Eureka, CA, USA.
- Sawyer, J. O., and T. Keeler-Wolf. 1995. A Manual of California Vegetation. California Native Plant Society, Sacramento, CA, USA.
- Spiegel, L., B. Cypher, and T. Dao. 1996. Diet of the San Joaquin kit fox (Vulpes macrotis mutica) at three sites in western Kern County, California. Pages 39–52 in L. Spiegel, editor. Studies of San Joaquin Kit Fox in Undeveloped and Oil Developed Areas. California Energy Commission, Sacramento, CA, USA.
- Stapp, P., A. McKenzie, D. M. Bucklin, R. A. Baldwin, and N. Quinn. 2025. Patterns of exposure of coyotes to anticoagulant rodenticides in California, USA. Journal of Wildlife Management 89:e22696.
- Thompson, B. 2014. Coyote and their movement in relation to resources in Tommy Thompson Park. Thesis, York University, Toronto, Ontario, Canada.
- White, L. A., and S. D. Gehrt. 2009. Coyote attacks on humans in the United States and Canada. Human Dimensions of Wildlife 14:419–432.
- Young, J. A., and C. G. Young. 1992. Seeds of Woody Plants in North America. Dioscorides Press, Portland, OR, USA.