

RESEARCH NOTE
Kristin Denryter1*, Thomas R. Stephenson2, and Kevin L. Monteith3
1 Alaska Department of Fish and Game, Wildlife Nutrition Lab, 1800 Glenn Highway, Suite 2, Palmer, AK 99645, USA ![]() https://orcid.org/0000-0003-0902-8213
https://orcid.org/0000-0003-0902-8213
2 California Department of Fish and Wildlife, Central Region, 787 North Main Street, Bishop, CA 93514, USA
![]() https://orcid.org/0000-0002-9412-8309
https://orcid.org/0000-0002-9412-8309
3 University of Wyoming, Department of Zoology and Physiology, Bim Kendall House, 804 East Fremont Street, Laramie, WY 82072, USA ![]() https://orcid.org/0000-0003-4834-5465
https://orcid.org/0000-0003-4834-5465
* Corresponding Author: kristin.denryter@alaska.gov
Published 28 August 2025 • doi.org/10.51492/cfwj.111.16
Key words: bomb calorimeter, digestible dry matter, digestible energy, digestibility, forage quality,gross energy, in vitro dry matter digestibility, IVDMD, multiplier effect, Ovis canadensis sierrae, sequential fiber analysis
| Citation: Denryter, K., T. R. Stephenson, and K. L. Monteith. 2025. Evaluating assumptions about gross energy in estimating nutritional value of forage: a case study using forages of Sierra Nevada bighorn sheep. California Fish and Wildlife Journal 111:e16. |
| Editor: Carly White, North Central Region |
| Submitted: 2 March 2025; Accepted: 30 June 2025 |
| Copyright: ©2025, Denryeter et al. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Competing Interests: The authors have not declared any competing interests. |
Nutrition is a basic requirement for biological life and sets the upper potential for wildlife population productivity (Morton and Cheathum 1946; Severinghaus 1951; Taber 1953; Cheathum and Severinghaus 1961; Julander et al. 1961; Skogland 1986; Monteith et al. 2014; Stephenson et al. 2020). For herbivores, nutritional resources often are evaluated in terms of food quality, measured as digestible energy (Parker et al. 1999; Tollefson et al. 2011; Cook et al. 2016)—the product of gross energy (measured as the amount of heat produced during combustion) and digestibility (the proportion of the food item that can be digested by the animal and not lost to feces). Given the time and money associated with assays to determine digestible energy, researchers are interested in alternatives for determining forage quality. Some researchers have used gross energy to index forage quality; however, not all gross energy is available to the animal (Robbins 1993). Others have assumed that variation in gross energy is small enough to be essentially inconsequential or have tried to eliminate the need to evaluate gross energy altogether by developing an equation for predicting digestible energy from digestibility (Spalinger et al. 2010). The broader applicability of assumptions about and relations among gross energy, dry matter digestibility, and digestible energy have not been evaluated, but they may represent oversimplifications of important phenomena. For example, although Robbins (1993) indicated that gross energy content of plant tissues was uniform, he also pointed out that energy content varies by plant part, growth form, season, plant type, and more. Hence, assumptions about gross energy, dry matter digestibility, and digestible energy warrant further exploration.
Differences in gross energy may become particularly important for foraging and nutrition or energy-balance studies where energy content of foods is multiplied up to some level of daily or seasonal intake. In these studies, food quality is multiplied over thousands of bites per day and millions of bites per season. Compounding errors in these types of studies therefore have the potential to result in significant multiplier effects (sensu Blaxter et al. 1961; White 1983). Multiplier effects relative to digestibility and digestible energy content of food have been reported previously for reindeer (Rangifer tarandus; Cebrian et al. 2008) and elk (Cervus elaphus; Cook et al. 2004). To our knowledge, compounding effects of variable gross energy have not been addressed in the literature but given that gross energy is a component of digestible energy, and that these can be scaled up to daily or seasonal intakes, the potential for important multiplier effects exists.
Climate in the Sierra Nevada consisted of dry summers (May–Sept) with little precipitation, save for afternoon thunderstorms with little accumulation, and wet winters, with most precipitation in the form of snow (Nov–Apr; Wehausen, 1980). Rugged topography and xeric vegetation communities (e.g., Great Basin sagebrush-bitterbrush scrub, pinyon-juniper woodlands) characterized the eastern slope of the Sierra Nevada. Mesic subalpine forest, subalpine meadows, and alpine meadows occur at higher elevations (>2,500 m; Wehausen 1980; Johnson et al. 2010). During summer, Sierra bighorn occupy habitats at moderate to high elevations (>3,000 m; Spitz 2015; Denryter et al. 2021b).
We collected samples of plant species available to Sierra Nevada bighorn (Ovis canadensis sierrae) sheep during June–August 2018, at peak quality as snowmelt progressed up in elevation, as part of a range assessment study. Typically, we identified plant samples to genus or species, but when we were unable to do so, we identified plants to family. Plant samples were collected separately by plant part (e.g., leaves, flowers, Table A1) and placed in plastic freezer bags kept cool with snow or cold water bottles (to stop plant metabolism and associated sample degradation) until we could hike out and transfer samples to a freezer. We submitted plant samples to the Washington State University Wildlife Habitat and Nutrition Lab where they were freeze dried, ground in a Wiley mill through a 1-mm mesh screen, and analyzed for gross energy (via bomb calorimetry in a C500 bomb calorimeter; Ika Works Inc., Wilmington, NC) and dry matter digestibility via sequential fiber analysis in an ANKOM fiber analyzer200/200® (ANKOM Technology, Fairport, NY) following published protocols from Ankom (Goering and Van Soest 1970; Ankom Technology n.d., 2017a, 2017b, 2022). We determined dry matter digestibility of forages using summative equations, assuming a value of zero for tannins (recognizing that forbs and shrubs contain some tannins that reduce dry matter digestibility; Robbins et al. 1987a,b). To ensure results most accurately related to animal performance, we used adjustments of Cook et al. (2022) because of a shift in results of lab assays after 2012 that biased estimates of digestibility for less-digestible forages, resulting in an overestimation of their digestibility. That work validated results from years of forage samples with nutritional requirements of animals and animal performance to illustrate the bias in samples assayed after 2012. To estimate digestible energy, we multiplied gross energy with dry matter digestibility. We ran single samples for each assay and all results are reported on a dry matter basis. Digestibility equations were developed for cervids, but comparable equations are not available for bovids, and these equations have recently become popularized for use in bighorn sheep (Wagler et al. 2023; Bilodeau-Hussey et al. 2025).
All statistical analyses were conducted in Stata 14 (StataCorp, College Station, TX). We used linear regression, ensuring all data met assumptions for linear regression first, to evaluate relations between gross energy, dry matter digestibility, and digestible energy for all samples pooled together and separately by forage class. Forage classes were deciduous shrubs, evergreen shrubs, forbs (which included flowers and ferns because of small sample size in those classes), and graminoids (i.e., grasses, sedges, rushes).
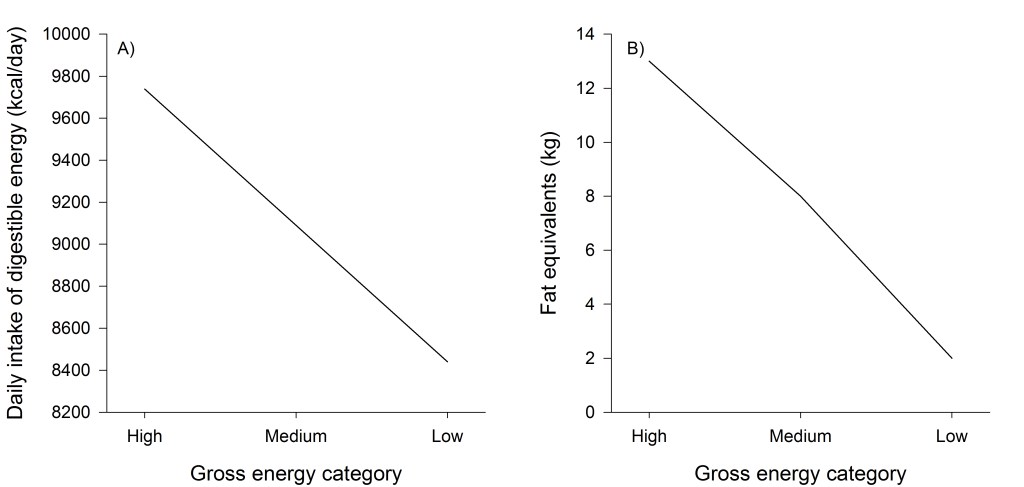
We used simple arithmetic models to illustrate how different levels of gross energy affected digestible energy content of forages, pooled by forage class due to small sample size for individual species, across 3 levels of gross energy. Levels for gross energy were high (mean plus 1 standard deviation), medium (mean value), and low (mean minus 1 standard deviation). We assumed a moderate level of dry matter digestibility (56.5%; weighted average by forage class). Using these values, we calculated the difference in digestible energy content of forages as gross energy changed. For simplicity, we calculated the difference in daily intake of digestible energy assuming a constant intake rate of 3.8 g/min for 14 h/day, while recognizing that changes in forage quality will affect digestive retention times, passage rates, and therefore intake rates (Mertens 1973; Allen and Mertens 1988; Minson and Wilson 1994). To evaluate seasonal energy balances, we took the difference between daily intakes of digestible energy and daily requirements for digestible energy over 120 days, however, this represents a best-case scenario as digestibility declines over time, Sierra bighorn may not eat all plants they encounter with equal probability, and forage biomass is depleted across the season (Cook et al. 2016; Denryter et al. 2022b; Johnson et al. 2022). Our model is intended for heuristic purposes only and not to reflect the complicated and dynamic foraging behavior of bighorn sheep that changes across the summer season. We assumed daily digestible energy requirements for lactation were ~8,218 kcal/day (~9,250 kcal/day minus ~1,032 kcal/day for tissue synthesis; Denryter et al. 2021a). We assumed the difference in digestible energy intake and requirements was available for use as fat and for simplicity assumed 65% efficiency of converting digestible energy to fat (Agricultural Research Council 1980; Boertje 1985). We converted energy to fat using a conversion of 1 kg of fat = 9,000 kcal (1 g = 9 kcal).
We collected a total of n = 70 plant samples for forage quality assays. For all samples, the mean (± SD) gross energy was 4.83 ± 0.36 kcal/g, the mean dry matter digestibility was 59.0 ± 6.2%, and the mean digestible energy was 2.85 ± 0.33 kcal/g; however, number of samples of each forage class were not equal and hence we also calculated a weighted mean. We calculated the weighted mean using mean values of each forage class multiplied by the proportion of samples in each forage class. Mean (± SD) gross energy, dry matter digestibility, and digestible energy content of forages when forage classes were given equal weight this way were 5.04 ± 0.36 kcal/g, 56.5 ± 6.75%, and 2.83 ± 0.19 kcal/g, respectively. Therefore, our values for high, medium, and low gross energy were 5.40 kcal/g, 5.04 kcal/g, and 4.68 kcal/g, respectively.
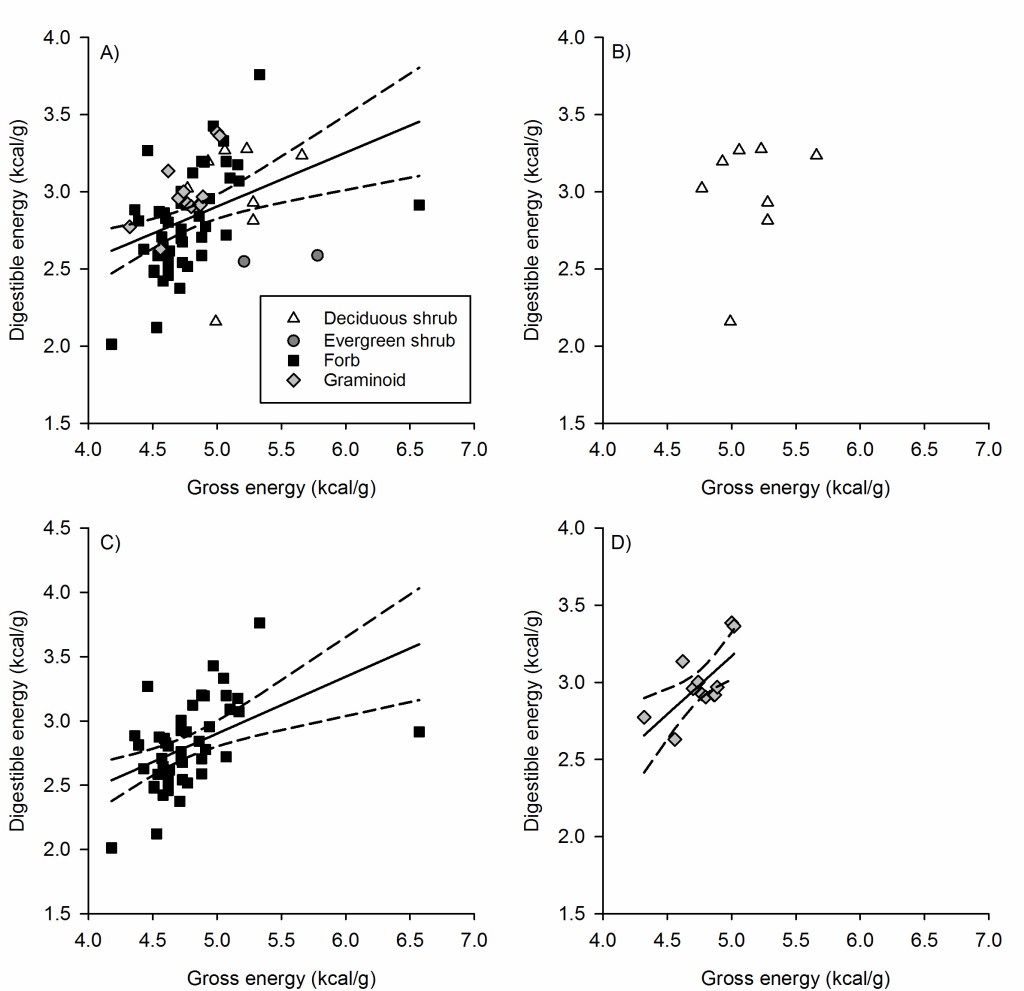
Across all samples, which were collected during late June–early August as snowmelt progressed representing peak quality (Table A1), the highest gross energy (6.57 kcal/g) occurred in a forb of the Brassicaceae family that we were not able to identify to species and the lowest gross energy occurred in the forb Monardella odoratissima (4.18 kcal/g) (Fig. 1). The highest dry matter digestibility occurred in the forb Sedum integrifolium (73.3%), and the lowest dry matter digestibility occurred in the deciduous (dwarf) shrub Salix arctica (43.3%). For digestible energy, the highest value occurred in the forb Ivesia pygmaea (3.76 kcal/g) and the lowest value in the forb Monardella odoratissima (2.01 kcal/g). Results by species are provided in Table A1.

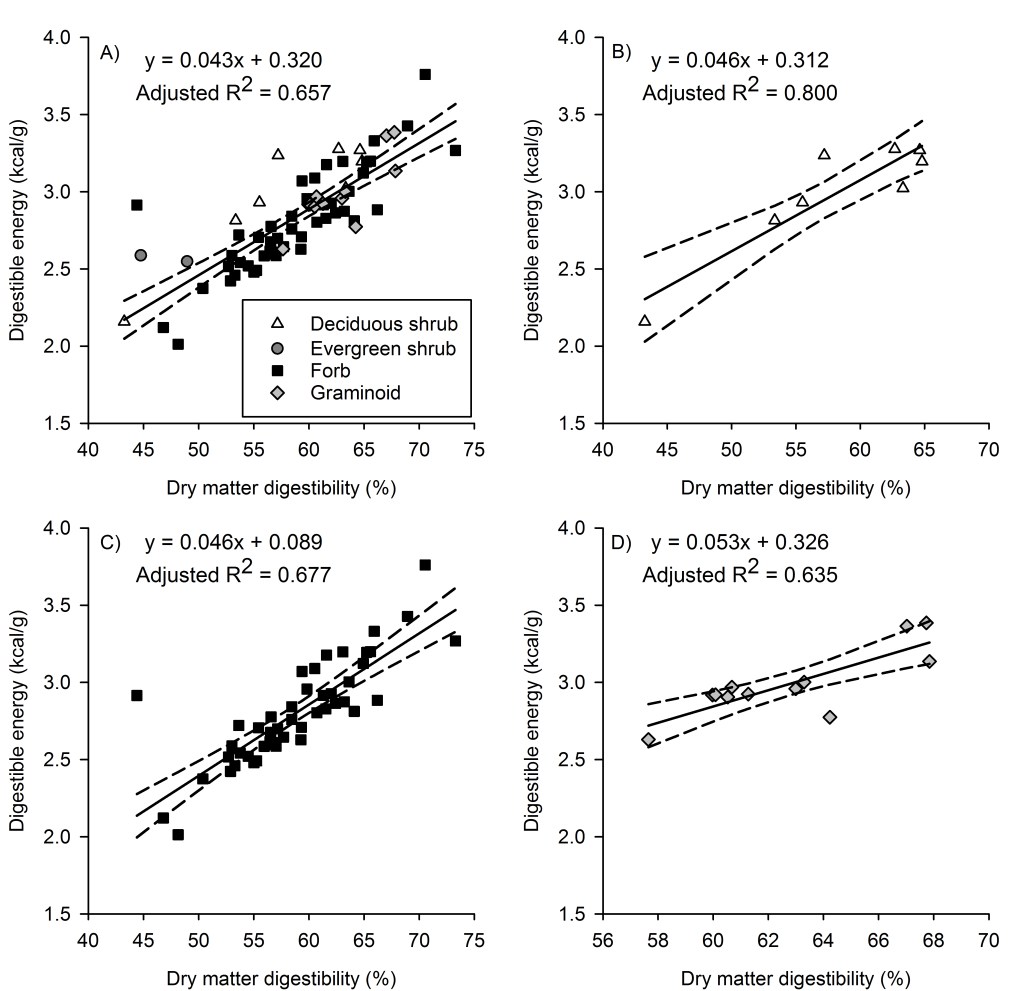
Dry matter digestibility was not a function of gross energy for all species combined (P = 0.078), deciduous shrubs (P = 0.652), forbs (P = 0.538), or graminoids (P = 0.672; Appendix A). We did not analyze evergreen shrubs separately owing to small sample size (n = 2). Digestible energy was a function of gross energy for all species combined (P = 0.001, F1,68 = 12.05, r2 = 0.151, adj r2 = 0.138), forbs (P < 0.001, F1,46 = 13.48, r2 = 0.227, adj r2 = 0.210), and graminoids (P = 0.015, F1,10 = 8.68, r2 = 0.565, adj r2 = 0.411), but not for deciduous shrubs (P = 0.579; Fig. 2). Digestible energy content was a function of dry matter digestibility for all species combined (P < 0.001, F1,68 = 133.35, r2 = 0.662, adj r2 = 0.657), for deciduous shrubs (P = 0.002, F1,6 = 133.35, r2 = 0.829, adj r2 = 0.800), for forbs (P < 0.001, F1,46 = 99.50, r2 = 0.683, adj r2 = 0.677), and for graminoids (P = 0.001, F1,10 = 20.16, r2 = 0.668, adj r2 = 0.635; Fig. 3).
The difference between daily intake at high levels of gross energy and low levels of gross energy was consistent, with high gross energy resulting in ~15% greater daily intake than low gross energy (Fig. 4). Differences in gross energy scaled up across the season to result in substantial differences in the amount of energy available for fat accretion, with high gross energy potentially allowing for accretion of 13 kg worth of fat compared to 8 kg at moderate gross energy and only 2 kg at low gross energy (Fig 4)—differences that could have significant implications to survival (Denryter et al. 2022a).
Quantifying the quality of available forage resources is a primary way through which researchers evaluate the nutritional value of habitats and their potential to support populations. Critical to these efforts is the application of appropriate techniques and metrics that accurately represent the nutritional value of forages to the forager of interest. Although gross energy is not an informative nutritional currency by itself, because not all gross energy is available to the animal for utilization (Robbins 1993), it is a key part of estimating nutritionally relevant currencies, including digestible energy. Our work demonstrates that: (1) gross energy can be quite variable within and among plant taxa (Table A1), (2) relations between dry matter digestibility and digestible energy are variable and estimating gross energy can help improve estimates of digestible energy, (3) gross energy alone cannot be used to reliably predict dry matter digestibility or digestible energy, and (4) small differences in gross energy can scale up to have large multiplier effects. Collectively, our analyses demonstrate that accounting for variable gross energy, in addition to dry matter digestibility, continues to be an important part of nutritional ecology research. We also caution that not accounting for variable gross energy could result in erroneous conclusions. That is not to say that all projects need gross energy to be successful, but that researchers should recognize where they may need to determine specific values (e.g., where multiplier effects may be important, such as when scaling to daily or seasonal intake to evaluate whether animals meet energy intake requirements) versus using published values (e.g., evaluating whether forages meet digestibility requirements). However, if researchers must choose one or the other because of budgetary limitations, dry matter digestibility should be prioritized over gross energy because of its greater ability to explain variation in digestible energy.
Gross energy of forages available to Sierra bighorn during summer was such that the largest values (6.6 kcal/g) were 57% greater than the smallest values (4.2 kcal/g), suggesting numerically small differences in gross energy were, in fact, proportionately large. Our models demonstrated that small differences in gross energy scaled up to have large multiplier effects on seasonal energy balance. A difference in gross energy of only 0.72 kcal/g (the difference between high and low gross energy simulations) altered digestible energy intake by 1,453 kcal/day, which over 120 days accumulated to a difference of the equivalent caloric value of ~11 kg of body fat between high and low gross energy. This heuristic example illustrates how small differences can scale up dramatically but should not be construed to reflect the true foraging behavior of Sierra bighorn—that would require a much more complicated modeling exercise that is beyond the scope of this article. The range of gross energy we measured, however, was more than three times the value we used in our model, which implies that differences in nutritional outcomes like body fat equivalents could be even greater than what we estimated. The fact that even a relatively small difference of <1 kcal/g of gross energy can result in substantial effects on energy available for fat accretion highlights the importance of determining gross energy of forages. This is particularly true for studies relating nutritional values of forages to nutritional requirements of animals to determine whether energy requirements can be met by available food supplies.
Although other authors have suggested that predicting digestible energy from dry matter digestibility was suitable for nutrition studies, our results contradict that assertion. While dry matter digestibility explained ~91% of variation in digestible energy in moose forages (Spalinger et al. 2010) and thereby suggested the need to estimate gross energy was superfluous, our analyses demonstrated that dry matter digestibility explained only ~66–83% of the variation in digestible energy content of bighorn forages. Given that our sensitivity analyses illustrated large cumulative impacts of small changes in gross energy, we conclude that predicting digestible energy content of forages from dry matter digestibility alone is less suitable for studies where compounding error is important. If economic constraints preclude determination of gross energy and dry matter digestibility, researchers will be better served by determining dry matter digestibility to understand forage quality, as indicated by better (but imperfect) relations between dry matter digestibility and digestible energy content of forages.
Our results also demonstrated that gross energy alone was not a suitable metric for evaluating forage quality. Animals do not have nutritional requirements for gross energy, because not all energy contained in a food item is available to the animal; some is lost to digestive and metabolic inefficiencies (Robbins 1993).For gross energy to be a suitable metric for evaluating forage quality, it would need to correlate with metrics that reflect the nutritional value of forages, such as dry matter digestibility or digestible energy, and explain a large amount of variation in digestible energy. However, gross energy content did not predict dry matter digestibility reliably (Fig. A1). Although a significant relation existed between gross energy and digestible energy for forbs, graminoids, and all species combined, it generally explained only 15–47% of variation in digestible energy for those species and hence was a poor to marginal predictor of digestible energy. Further illustrating why gross energy has limited utility as an independent index of nutritional value of forage is the fact that evergreen shrubs had the highest gross energy of all forage classes. Had we used gross energy to index forage quality, we would have erroneously concluded that evergreen shrubs were the highest-quality forage because their gross energy was 6% higher than for deciduous shrubs and 13–14% higher for graminoids and forbs. In reality, evergreen shrubs were the lowest in forage quality, with digestible energy averaging ~11% lower than for any other forage class; their higher gross energy resulted from high content of energy-dense volatile oils (Robbins 1993).
Accurately estimating forage quality is essential in understanding the nutritional value of forages and potential for those forages to support wild herbivores. While our findings demonstrate that gross energy is not a suitable metric of forage quality, they also highlight the importance of accurately estimating gross energy as an input to digestible energy. Gross energy is not uniform among plants, even within the same species, and even small differences can scale up to have large multiplier effects on seasonal energy balances. Animals do not have requirements for dry matter digestibility, but it generally correlates with digestible energy and can constrain digestion and intake rates at low levels (Mertens 1973; Bergman et al. 2001). Further, dry matter digestibility had more explanatory power than gross energy as a determinant of digestible energy content of forages. Animals do have requirements for digestible energy, and it integrates gross energy and dry matter digestibility, making it a superior and quantitative metric for evaluating forage quality. Collectively, our findings underscore the importance of accurately estimating gross energy, in addition to dry matter digestibility, in studies of forage quality. Failing to account for differences in gross energy may lead to erroneous conclusions in the nutritional value of forages and potential consequences to the conservation and management of herbivores relying on them.
Acknowledgments
We are grateful to T. Calfee, D. German, L. Greene, B. Hatfield, C. Massing, E. Otto, E. Siemion, A. Sturgill, J. Wehausen, J. Weissman, and J. White (Leary) for field assistance.
Literature Cited
- Agricultural Research Council. 1980. The Nutrient Requirements of Ruminant Livestock. Agricultural Research Council. Commonwealth Agricultural Bureau, Slough, England.
- Allen, M. S., and D. R. Mertens. 1988. Evaluating constraints on fiber digestion by rumen microbes. Journal of Nutrition 118:261–270.
- Ankom Technology. 2017a. Acid Detergent Fiber in Feeds Filter Bag Technique (for A200 and A200I) ADF Method Method 5. Ankom Technology: 1–2. Available from: https://www.ankom.com/analytical-methods-support/fiber-analyzer-a2000 (Accessed 30 June 2025)
- Ankom Technology. 2017b. Neutral Detergent Fiber in Feeds. Filter Bag Technique (For A200,A200I) – NDF Method 6. ANKOM Technology. Available from: http://www.ankom.com/09_procedures/NDF_081606_A200.pdf (Accessed 30 June 2025)
- Ankom Technology. 2022. Method 8 – determining Acid Detergent Lignin in beakers. Ankom Technology. Available from: https://www.ankom.com/sites/default/files/document-files/Method_8_Lignin_in_beakers.pdf (Accessed 30 June 2025)
- Ankom Technology. (n.d.). Ashing procedure Service Procedure 034. Available from: https://www.ankom.com/sites/default/files/document-files/AS034_Ashing_Procedure.pdf (Accessed 30 June 2025)
- Bergman, C. M., J. M. Fryxell, C. C. Gates, and D. Fortin. 2001. Ungulate foraging strategies: energy maximizing or time minimizing? Journal of Animal Ecology 70:289–300.
- Bilodeau-Hussey, N. M., K. S. Huggler, E. F. Cassirer, H. Miyasaki, M. A. Hurley, L. A. Shipley, and R. A. Long. 2025. Effects of maternal condition, disease status, and behavior on survival of juvenile bighorn sheep. Journal of Wildlife Management 89:e22721.
- Blaxter, K. L., F. W. Wainman, and R. S. Wilson. 1961. The regulation of food intake by sheep. Animal Production 3:51–61.
- Boertje, R. D. 1985. An energy model for adult female caribou of the Denali Herd, Alaska. Journal of Range Management 38:468–473.
- Cebrian, M. R., K. Kielland, and G. Finstad. 2008. Forage quality and reindeer productivity: multiplier effects amplified by climate change. Arctic, Antarctic, and Alpine Research 40:48–54.
- Cheathum, E .L., and C. W. Severinghaus. 1961. Variations in fertility of white-tailed deer related to range conditions. Transactions of the North American Wildlife Conference 15:170–189.
- Cook, J. G., R. C. Cook, R. W. Davis, and L.L. Irwin. 2016. Nutritional ecology of elk during summer and autumn in the Pacific Northwest. Wildlife Monographs 195:1–81.
- Cook, J. G., B. K. Johnson, R. C. Cook, R. A. Riggs, L. D. Bryant, and L. L. Irwin. 2004. Effects of summer-autumn nutrition and parturition date on reproduction and survival of elk. Wildlife Monographs 155:1–61.
- Cook, R. C., L. A. Shipley, J. G. Cook, M. J. Camp, D. S. Monzingo, S. L. Robatcek, S. L. Berry, I. T. Hull, W. L. Myers, K. Denryter, and R. A. Long. 2022. Sequential detergent fiber assay results used for nutritional ecology research: evidence of bias since 2012. Wildlife Society Bulletin 46:e1348.
- Denryter, K., M. M. Conner, T. R. Stephenson, D. W. German, and K. L. Monteith. 2022a. Survival of the fattest: how body fat and migration influence survival in highly seasonal environments. Functional Ecology 36:2569–2579.
- Denryter, K., R. C. Cook, J. G. Cook, and K. L. Parker. 2022b. Animal-defined resources reveal nutritional inadequacies for woodland caribou during summer in northeastern British Columbia. Journal of Wildlife Management 86:e22161.
- Denryter, K., D. W. German, T. R. Stephenson, and K. L. Monteith. 2021a. State- and context-dependent applications of an energetics model in free-ranging bighorn sheep. Ecological Modelling 440:109349.
- Denryter, K., T. R. Stephenson, and K. L. Monteith. 2021b. Broadening the migratory portfolio of altitudinal migrants. Ecology 102:e03321.
- Goering, H. K., and P. J. Van Soest. 1970. Forage fiber analysis. Agriculture Handbook No. 379. U.S. Department of Agriculture, Agriculture Research Service, Washington, D.C., USA.
- Johnson, H. E., L. S. Mills, T. R. Stephenson, and J. D. Wehausen. 2010. Population-specific vital rate contributions influence management of an endangered ungulate. Ecological Applications 20:1753–1765.
- Johnson, H. E., E. A. Lenart, D. D. Gustine, L. A. Adams, and P. S. Barboza. 2022. Survival and reproduction in Arctic caribou are associated with summer forage and insect harassment. Frontiers in Ecology and Evolution 10:899585.
- Julander, O., W. L. Robinette, and D. A. Jones. 1961. Relation of summer range condition to mule deer herd productivity. Journal of Wildlife Management 25:54–60.
- Mertens, D. R. 1973. Application of Theoretical Mathematical Models to Cell Wall Digestion and Forage Intake in Ruminants. Cornelly University, Ithaca, NY, USA.
- Minson, D. J., and J. R. Wilson. 1994. Prediction of intake as an element of forage quality. Pages 553–563 in G. C. Fahey, editor. Forage Quality, Evaluation and Utilization. American Society of Agronomy, Madison, WI, USA.
- Monteith, K. L., V. C. Bleich, T. R. Stephenson, B. M. Pierce, M. M. Conner, J. G. Kie, and R. T. Bowyer. 2014. Life-history characteristics of mule deer: effects of nutrition in a variable environment. Wildlife Monographs 186:1–61.
- Morton, G., and E. L. Cheathum. 1946. Regional differences in breeding potential of white-tailed deer in New York. Journal of Wildlife Management 10:242–248.
- Parker, K. L., M. P. Gillingham, T. A. Hanley, and C. T. Robbins. 1999. Energy and protein balance of free-ranging black-tailed deer in a natural forest environment. Wildlife Monographs 143:1–48.
- Robbins, C. T. 1993. Wildlife Feeding and Nutrition. 2nd edition. Academic Press Inc., San Diego, CA, USA.
- Robbins, C. T., T. A. Hanley, A. E. Hagerman, O. Hjeljord, D. L. Baker, C. C. Schwartz, and W.W. Mautz. 1987a. Role of tannins in defending plants against ruminants: reduction in protein availability. Ecology 68:98–107.
- Robbins, C. T., S. Mole, A. E. Hagerman, and T. A. Hanley. 1987b. Role of tannins in defending plants against ruminants: reduction in dry matter digestion. Ecology 68:1606–1615.
- Severinghaus, C. W. 1951. A study of productivity and mortality of coralled deer. Journal of Wildlife Management 15:73–80.
- Skogland, T. 1986. Density dependent food limitation and maximal production in wild reindeer herds. Journal of Wildlife Management 50:314–319.
- Spalinger, D. E., W. B. Collins, T. A. Hanley, N. E. Cassara, and A. M. Carnahan. 2010. The impact of tannins on protein, dry matter, and energy digestion in moose (Alces alces). Canadian Journal of Zoology 88:977–987.
- Spitz, D. B. 2015. Does migration matter? Cause and consequences of migratory behavior in Sierra Nevada bighorn sheep. Dissertation, University of Montana, Missoula, MT, USA.
- Stephenson, T. R., D. W. German, E. F. Cassirer, D. P. Walsh, M. E. Blum, M. Cox, K. M. Stewart, and K. L. Monteith. 2020. Linking population performance to nutritional condition in an alpine ungulate. Journal of Mammalogy 101:1244–1256.
- Taber, R. D. 1953. Studies of the black-tailed deer reproduction on three chaparral cover types. California Fish and Game 39:177–186.
- Tollefson, T. N., L. A. Shipley, W. L. Myers, and N. Dasgupta. 2011. Forage quality’s influence on mule deer fawns. Journal of Wildlife Management 75:919–928.
- Wagler, B. L., R. A. Smiley, A. B. Courtemanch, D. Lutz, D. McWhirter, D. Brimeyer, P. Hnilicka, T. J. Robinson, and K. L. Monteith. 2023. Implications of forage quality for population recovery of bighorn sheep following a pneumonia epizootic. Journal of Wildlife Management 87:e22452.
- Wehausen, J. D. 1980. Sierra Nevada bighorn sheep: history and population ecology. Dissertation, University of Michigan, Ann Arbor, MI, USA.
- White, R. G. 1983. Foraging patterns and their multiplier effects on productivity of northern ungulates. Oikos 40:377–384.