

SHORT COMMUNICATION
William B. Ludt1*, Michelle C. Horeczko2, Todd R. Clardy1, and Benjamin W. Frable3
1 Natural History Museum of Los Angeles County, Department of Ichthyology, 900 Exposition Boulevard, Los Angeles, CA 90007, USA![]() https://orcid.org/0000-0002-0599-9699 (WBL)
https://orcid.org/0000-0002-0599-9699 (WBL)![]() https://orcid.org/0000-0002-7925-8789 (TRC)
https://orcid.org/0000-0002-7925-8789 (TRC)
2 California Department of Fish and Wildlife, Marine Region, 2451 Signal Street, Berth 57, San Pedro, CA 90731, USA
3 University of California, San Diego, Scripps Institution of Oceanography, 9500 Gilman Drive, La Jolla, CA 92093, USA ![]() https://orcid.org/0000-0003-4525-0671
https://orcid.org/0000-0003-4525-0671
*Corresponding Author: wludt@nhm.org
Published 28 August 2025 • doi.org/10.51492/cfwj.111.15
Key words: anglerfish, Ceratioidei, deep-sea fish, Himantolophus sagamius, rarity
| Citation: Ludt, W. B., M. C. Horeczko, T. R. Clardy, and B. W. Frable. 2025 Recent beach strandings of the Pacific footballfish on the Pacific coast of the United States. California Fish and Wildlife Journal 111:e15. |
| Editor: Helen Killeen, Marine Region |
| Submitted: 23 May 2025; Accepted: 30 June 2025 |
| Copyright: ©2025, Ludt et al. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Competing Interests: The authors have not declared any competing interests. |
Oceanic anglerfishes of the suborder Ceratioidei (sensu Pietsch 2009) are charismatic deep-sea fishes that have captured the public’s imagination, appearing in movies, television shows, and popular culture. Living at inaccessible depths with many species displaying large mouths, sharp teeth, and bioluminescent features, it’s easy to see why they are so recognizable. While most people are familiar with anglerfishes generally, many may not realize how diverse these fishes are, both in terms of species diversity but also body shape and life history. The suborder comprises 179 species (Fricke et al. 2025) that occur worldwide in the depths of tropical and temperate oceans, around 20 of which occur in coastal California (Love et al. 2021). Some species are relatively common and well-represented in the literature and museum collections, and several species have been documented in-situ giving us a glimpse into their behaviors and life history (Luck and Pietsch 2008; Lundsten et al. 2012; Stewart et al. 2023). However, others are exceedingly rare.
One such species is the Pacific Footballfish, Himantolophus sagamius (Tanaka 1918). Himantolophus is the only genus in the family Himantolophidae, which comprises 23 species (Fricke et al. 2025), including two species off California: H. nigricornis (Bertelsen and Krefft 1988) and H. sagamius (Love and Passarelli 2020). When this family was revised by Bertelsen and Krefft (1988), a mere 25 metamorphosed females of the Pacific Footballfishwere known to have been preserved, including two caught off California (Lea 1988). At that time two live observations of this rare species were known: one from a specimen that washed ashore in Japan and was kept in an aquarium for eight days (Haneda 1968), and one from Monterey, California, that was kept alive for 65 hours at the Monterey Bay Aquarium (Lea 1988). Since the revision, new observations have occurred and several specimens have been collected. Although this species is exceedingly rare throughout its range, an unusual number of specimens have washed ashore along the west coast of the United States in the past five years. Here we detail these new records in the context of all known specimens of this species to determine if these new records can reveal additional insights into this rare species.
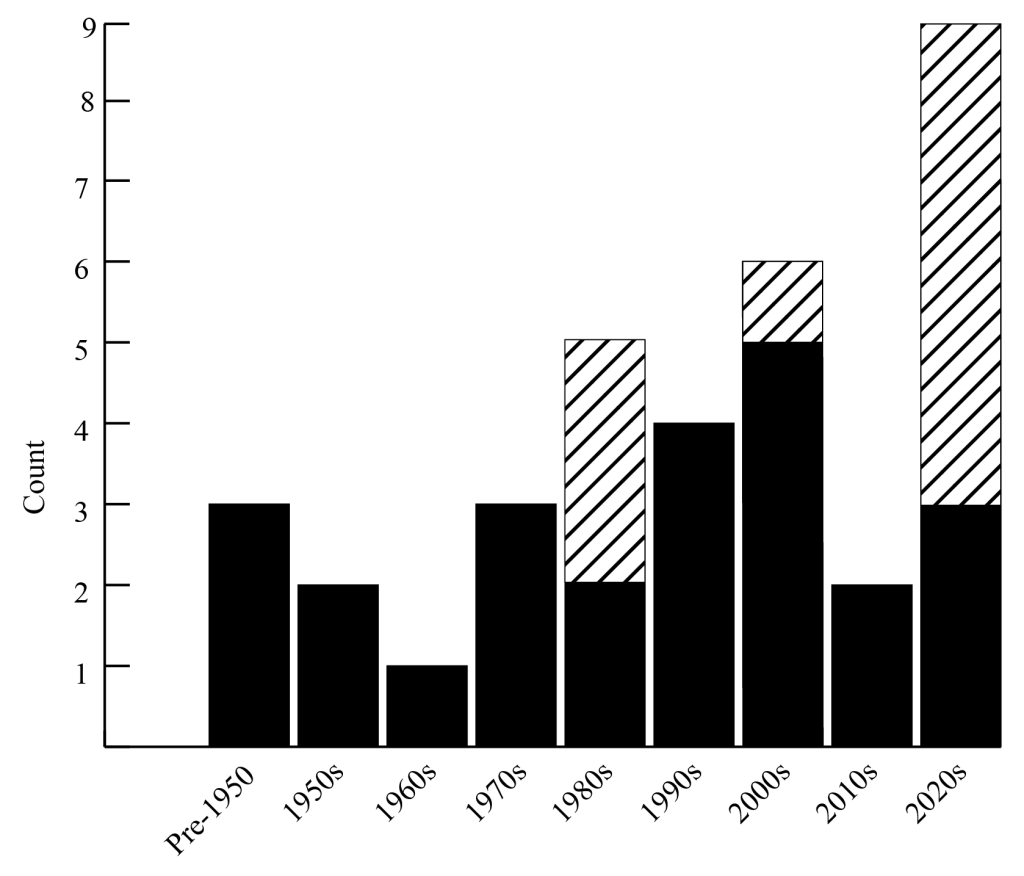
Information regarding all known specimens of this single species was gathered either from online repositories, such as Fishnet2.net, or by directly contacting museum collections. Data from recent California and Oregon specimens were gathered by direct observation of collected specimens or from photographs of specimens that were not collected, which were identified by diagnostic illicial and escal characters (Pietsch 2009). In total, 41 mature female individuals of this species have been recorded since 1907, seven of which were either lost or not preserved (Table 1). No males of this species have been described. Nineteen of the records come from the northwest Pacific (16 from Japan, three from Russia), 15 from the northeast Pacific (11 from California, one from Oregon, one in the northern Pacific, two off Hawai’i), three from the southwest Pacific (all from Indonesia), and four from the southeast Pacific (two from the Galapagos, one from Chile, one from Peru). Three specimens have been reported from the vicinity of Palau, but their identification is tentative due to their small size (14, 32, and 45 mm standard length (SL), respectively) and therefore they are not included in the counts here. While data is not available for every individual documented, specimens range in size from 32–380 mm SL and have been recovered from a variety of depths. Eleven specimens were beach strandings, one was caught in a gill net at approximately 30 m, and the majority were caught in various types of midwater trawls at depths ranging from 60 to >1000 m. All but three specimens were recorded after 1950, with 21 records since 2000 (Fig. 1). This highlights two trends: increased activity in deep-sea fishing/sampling and increased access to photographic devices such as cameras and smartphones.
Table 1. All known specimens and observations of the Pacific Footballfish along with their date, location, depth, standard length (SL), and comments. All museum acronyms follow Sabaj 2020. B&K represents Bertelsen and Krefft 1988.
| No. | Catalog No. | Date | Country/ Location | Lat/Long | Depth (m) | SL (mm) | Comments |
| 1 | SCMT 8201 (holotype) | Prior to 1918 | Japan | 35ºN 139º30’E | ND | 200 | Specimen lost. Coordinates estimated in B&K. |
| 2 | SCMT 8460 | Prior to 1918 | Japan | ND | ND | 274 | Specimen lost. |
| 3 | MCZ 29854 | 1 Feb 1907 | Japan | 34º51.55’N 139°20.69’E | 1280 | 231 | — |
| 4 | ZMH-ICH- 0100258a | 1954 | Ecuador (Galapagos) | ND | NA | 160 | Reported in B&K as recovered from sperm whale stomach. |
| 5 | ZMH-ICH- 0100258b | 1954 | Ecuador (Galapagos) | ND | NA | 240 | Reported in B&K as recovered from sperm whale stomach. |
| 6 | HUMZ 17738 | Prior to 1960 | Japan | ND | ND | ND | Reported from Aomori Prefecture. Date estimate provided by F. Tashiro, HUMZ, personal communication. |
| 7 | Observation Only | 1968 | Japan | ND | 0 | 295 | Beach stranding kept in aquarium for 7 days. Disposed of after death. Published in Haneda 1968. |
| 8 | LACM 42698-1 | 22 Jun 1971 | USA (Hawai’i) | 21°20’N 158º20’W | 630– 670 | 37 | Caught with pelagic closing net. Tentative juvenile – Pietsch 2009. |
| 9 | SIO 72-13 | 25 Sep 1971 | North Pacific | 27°22.30’N 155º04’E | ND | 37 | Caught with midwater trawl. |
| 10 | LACM 36073-10 | 5 May 1975 | Indonesia | 5°1.5’S 130º04.7’E | 650– 1000 | 32 | Caught with midwater trawl. Tentative juvenile – Pietsch 2009. |
| 11 | ZMDVGU IV 7600/1014 | 4 Jan 1980 | Russia (Sakhalin Oblast) | 44º20’N 148º37’E | 1200 | 186 | Caught with otter trawl. Reported in Kharin 2006. |
| 12 | LACM 43760-1 | 18 Dec 1983 | USA (California) | 35°13’N 121º16’W | 613 | 111 | — |
| 13 | HUMZ 103697 | 30 Apr 1984 | Japan | ND | 800 | Caught with bottom trawl. | |
| 14 | CAS 57639 | 3 Jan 1985 | USA (California) | 36º47’N 122º01’W | 914– 969 | 180 | Midwater capture over center of Monterey Submarine Canyon. |
| 15 | Observation Only | Prior to 1988 | Japan | 35º45’N 141ºE | ND | 128 | Caught with bottom trawl. Specimen lost. Unknown date but reported as personal communcation from T. Abe in B&K. |
| 16 | CMA 1990.09.0001 | 14 Nov 1989 | USA (California) | 32º22’N 118º27’W | 30 | 372 | Caught with gill net. |
| 17 | MNHNC P. 6848 | 1991 | Chile | 29°57’S 71°22’W | ND | 111 | — |
| 18 | NSMT 59015 | 10 Jul 1996 | Japan | 40º43.4’N 143º10.6’E | 530– 558 | 73 | — |
| 19 | NSMT 58536 | 30 Jul 1996 | Japan | 39º0.2’N 143º29.7’E | 650– 679 | 69 | — |
| 20 | HUMZ 167672 | 1999 | Peru | ND | ND | 332.2 | Caught with midwater beam trawl. |
| 21 | SIO 02-2 | 15 Dec 2001 | USA (California) | 32º57.5’N 117º15.9’W | 0 | 380 | Beach stranding. |
| 22 | NSMT 69976 | 17 Aug 2004 | Japan | 40º30’N 170º20’E | 60– 110 | 223 | — |
| 23 | HUMZ 191925 | 13 Feb 2005 | USA (Hawai’i) | 13°34.32’N 158º46.32’W | ND | 296.2 | Caught with drift line. |
| 24 | HUMZ 194261 | 14 May 2005 | Indonesia | 8°35.04’S 110º43.02’E | 605– 607 | 43 | Caught with bottom trawl. |
| 25 | HUMZ 194599 | 16 Jul 2005 | Indonesia | 8°17.52’S 109º44.1’E | 612– 632 | 50.9 | Caught with bottom trawl. |
| 26 | NSMT 90520 | 1 Nov 2007 | Japan | 36º30.9’N 141º21.1’E | 899 | 41 | — |
| 27 | SIOM | Prior to 2009 | Russia (Sakhalin Oblast) | ND | ND | 45 | Caught with midwater trawl. Reported in Pietsch 2009. |
| 28 | IMBV | Prior to 2009 | Russia (Sakhalin Oblast) | ND | ND | 186 | Reported in Pietsch 2009. |
| 29 | HUMZ 211661 | 15 Dec 2010 | Japan | 26°46.62’N 141º57.9’E | 570– 730 | ND | Caught with midwater beam trawl. |
| 30 | KPM-NI 28001 | 16 Feb 2011 | Japan | 35°1.84’N 138º47.33’E | 0 | 146.8 | Beach stranding. |
| 31 | LACM 60082-1 | 5 Jul 2021 | USA (California) | 33º34.5’N 117º50.52’W | 0 | 295 | Beach stranding. |
| 32 | Observation Only | 13 Nov 2021 | USA (California) | 32°53.45’N 117º15.25’W | 0 | ND | Stranding at Black’s Beach, CA, USA. Specimen lost. |
| 33 | SIO 21-41 | 10 Dec 2021 | USA (California) | 33°02.03’N 117°17.51’W | 0 | 282 | Beach stranding. |
| 34 | FAKU 149078 | Prior to 2022 | Japan | ND | 1000 | ND | While collection date unknown, a tissue of this specimen is used in Kai et al. 2022. Reported from Miyagi Prefecture. |
| 35 | FAKU 149647 | 30 Oct 2022 | Japan | 36º34.24’N 141º21.42’E | 758 | 208.8 | Caught with otter trawl. |
| 36 | Observation Only | 17 Jun 2023 | USA (California) | 33º12’N 117º23.44’W | 0 | Stranding in Oceanside, CA, USA. Specimen lost. | |
| 37 | FAKU 150947 | 9 Oct 2023 | Japan | 40º13.74’N 142º15.54’E | 460 | 84.8 | Caught with otter trawl. |
| 38 | LACM 61593 | 13 Oct 2023 | USA (California) | 33º34.5’N 117º50.52’W | 0 | 310 | Beach stranding. |
| 39 | Observation Only | 18 May 2024 | USA (Oregon) | 45º53.46’N 123º57.9’W | 0 | ND | Stranding at Cannon Beach, OR, USA. Specimen lost. |
| 40 | SIO 25-10 | 12 Feb 2025 | USA (California) | 33º05.07’N 117º18.76’W | 0 | 230 | Beach stranding. |
| 41 | SIO 25-43 | 25 Jun 2025 | USA (California) | 33º15.45’N 117º26.27’W | 0 | 246 | Beach stranding found partially desiccated and damaged. |
Eleven specimens have been documented in California, eight of which washed ashore (with seven of those occurring within the last five years; Table 1). The first two California specimens, collected using midwater trawls in 1983 and 1985, are the smallest at 111 mm and 180 mm SL, respectively. The remaining beach strandings, and one specimen that was caught in a gill net at 30 m depth, are all 230 mm SL or larger. All California specimens have been documented south of Point Conception with the exception of one individual from Monterey Bay (CAS 57639) and one from Morro Bay (LACM 43760-1; Fig. 2). None have shown external damage consistent with predation prior to stranding, or any known cause of death that may have led to their beaching, and all were recovered with empty stomachs.

Perhaps the most variable external feature of Himantolophus anglerfishes is their bioluminescent esca (Pietsch 2009), and with so few specimens known it is important to document the variation that exists within each species as the amount of intraspecific and interspecific variation is still unknown (Kai et al. 2022). Of the recent material for the Pacific Footballfish, five were preserved and accessioned into museum collections (Fig. 3; two California specimens were lost, presumably to scavengers or to rising tides, and the fate of the Oregon specimen is unknown). The general morphology of the body, illicium and escal bulb of these specimens closely matches those outlined by Bertelsen and Krefft (1988), Lea (1988), Klepadlo et al. (2003), and Pietsch (2009). The esca has two distal appendages with bifurcating, blunt tips, each with a smaller side branch near the base. At the base of the distal appendage are four blunt lobes. There is a simple, unbranched anterior escal appendage, one simple or bifurcating posterior escal appendage, and a variable number of illicial appendages stemming from the illicium. Variation outside the range of previous publications is present in the number of dermal spines as well as the number and branching pattern of illicial appendages. Pietsch (2009) reported a range of 40–75 dermal spines on each side of the body in known specimens at the time. While some recent specimens fall within that range (SIO 21-41 and LACM 60082-1), one individual has 37 dermal spines on each side of its body (SIO 25-10), while another has 35 dermal spines on the left side of the body, and 37 on the right (LACM 61593). Unequal numbers of dermal spines were also reported by Klepadlo et al. (2003). Furthermore, Pietsch (2009) reported four to five dermal spines at the base of each pectoral fin, while we find as few as two in one individual (LACM 61593), and as many as six in another (LACM 60082-1). Regarding the illicium, Bertelsen and Krefft (1988) reported as few as two and as many as eight illicial appendages among the eleven specimens they examined, with some bifurcated and others simple. In the four most recent strandings, one has nine illicial appendages (two bifurcate and seven simple; LACM 61593), one has six (two bifurcate and four simple; LACM 60082-1), two have four (two bifurcate and two simple; SIO 21-41and SIO 25-10) and one has five (two bifurcate and three simple; SIO 25-43). The most recently collected specimen (SIO 25-43) was discovered desiccated and damaged at Marine Corps Base Camp Pendleton, San Diego County, making morphometric and meristic comparisons difficult, however, the IA appear intact without evidence of damage. Complete measurements of the esca and illicial appendages for the four recent specimens (excluding SIO 25-43) can be found in Table 2.
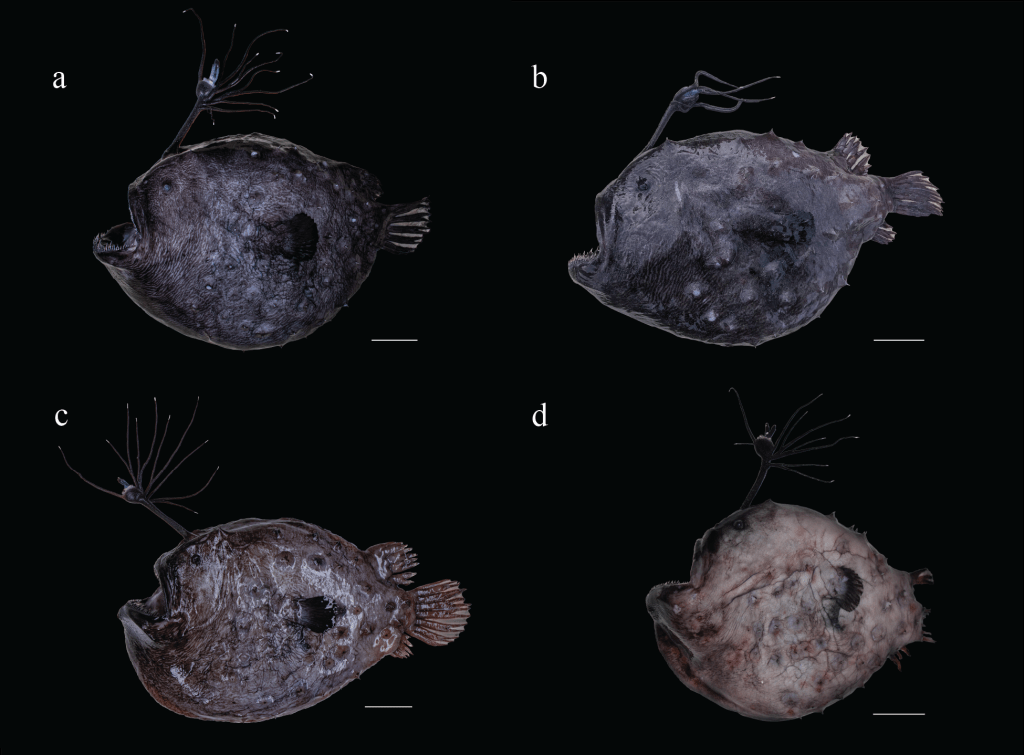
Table 2. Counts and measurements of four recent specimens that were catalogued into natural history museums. All measurements are in millimeters followed by percent of standard length (SL) in parentheses. DA – distal appendage, AA – anterior escal appendage, PA – posterior escal appendage, IA – illicial appendage. Illicial appendages abbreviated as documented by Klepadlo et al. (2003). Not included is SIO 25-43, as it was discovered dried and damaged, but with 5 intact IA and no evidence of others missing.
| Catalog Number | LACM 60082-1 | SIO 21-41 | LACM 61593 | SIO 25-10 |
| Year Cataloged | 2021 | 2021 | 2023 | 2025 |
| SL | 310 | 282 | 325 | 217 |
| Length of illicium | 97 (31.3) | 114.3 (40.5) | 98 (30.2) | 93.9 (43.3) |
| Diameter of escal bulb | 16 (5.7) | 14.2 (5.0) | 18 (5.5) | 12.4 (5.7) |
| Length of DA | 29 (9.4) | 17.3 (6.1) | 17 (5.2) | 14.8 (6.8) |
| Length of AA | 100 (32.3) | 100.9 (35.8) | 90 (27.7) | 59.4 (27.4) |
| Length of PA | 88 (28.4) | 103.5 (36.7) | 95 (29.2) | 70.2 (32.3) |
| Length of longest IA | 129 (41.6) | 119.6 (42.4) | 122 (37.5) | 88.8 (40.9) |
| No. of IA | 7 | 4 | 7 | 4 |
While little is still known about this species, it is clear that an increasing frequency of observations is becoming available, and every single specimen can teach us things not only about this species, but anglerfishes more generally (Ludt and Clardy 2022). One of the most puzzling things about this species is that a significant portion of California records are associated with individuals that have washed ashore, far from the depth they usually inhabit. No seasonal trends to specimen strandings are detected in this dataset, however these beached individuals are some of the largest specimens known, with the exception of one smaller individual (SIO 25-10). It is possible that these beached individuals are malnourished and nearing the end of their lives as a recent video shows a disoriented large female (presumably KPM-NI 28001) moving uncharacteristically three days prior to it washing ashore (H. Senou, Kanagawa Museum, pers. comm.). While there is no known cause for why this species occasionally washes ashore, additional specimens or observations may reveal important life history characteristics that explain this phenomenon.
These new records have already increased the known number of illicial appendages present on this species and updated our understanding of the range of dermal spines this species can have. They have also increased the known distribution of the Pacific Footballfish, as the 2024 Cannon Beach, Oregon, specimen represents the northern-most documented individual of this species. Additionally, no known males of this species have ever been reported and are awaiting description from future collections. While the range of this species is vast, occurring across the entire Pacific Ocean, the most common areas where it is reported are also some of the most densely populated: California and Japan. As such, we look forward to future specimens and the knowledge we can glean from them, and encourage beachgoers and fishermen to report unique fishes they encounter to their local fish and game office, natural history museum or university.
Acknowledgments
We thank the following people in no particular order for their assistance in gathering data about specimens: D. Catania (California Academy of Sciences), J. Luis Canto Hernandez (Chilean National Museum of Natural History), J. Hunt (Crystal Cove State Park), Y. Kai (Kyoto University), E. Karmovskaya (Institute of Oceanography, Russian Academy of Sciences), M. Nakae (Japan National Museum of Nature and Science), J. Passarelli (Cabrillo Marine Aquarium), T. Pietsch (University of Washington, retired), H. Senou (Kanagawa Prefecture Museum of Natural History), M. Schaadt (Cabrillo Marine Aquarium, retired), F. Tashiro (Hokkaido University Museum), J. Ugoretz (California Department of Fish and Wildlife), and staff at Crystal Cove State Park. Additionally, we thank D. Arcila, E. Ignatoff, E. McFarland, and J. Zhong (Scripps Institution of Oceanography), J. Hyde (Southwest Fisheries Science Center, National Oceanic and Atmospheric Administration), K. Carmody (MCB Camp Pendleton), and J. Coronel for assistance with recovering the San Diego specimens.
Literature Cited
- Bertelsen, E., and G. Krefft. 1988. The ceratioid family Himantolphidae (Pisces, Lophiiformes). Steenstrupia 14(2):9–89.
- Fricke, R., W. N. Eschmeyer, and J. D. Fong. 2025. Eschmeyer’s Catalog of Fishes: Genera/Species by Family/Subfamily. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (Accessed 30 Apr 2025)
- Haneda, Y. 1968. Observations on the luminescence of the deep-sea luminous anglerfish, Himantolophus groenlandicus. Science Report of the Yokosuka City Museum 14:1–9.
- Kai, Y., A. Otani, R. Misawa, B. W. Frable, and F. Tashiro. 2022. First Records of a Rare Deep-sea Anglerfish, Himantolophus azurlucens, from the Western North Pacific, with Comments on the DNA Barcodes of the genus (Lophiiformes: Himantolophidae). Species Diversity, 27(2):285–292. https://doi.org/10.12782/specdiv.27.285
- Kharin, V.E. 2006. Himantolophus sagamius (Himantolophidae), a new fish species for fauna of Russia. Journal of Ichthyology 46(3):274–275. https://doi.org/10.1134/S0032945206030088
- Klepadlo, C., P. A. Hastings, and R. H. Rosenblatt. 2003. Pacific Footballfish, Himantolophus sagamius (Tanaka) (Teleostei: Himantolophidae), found in the surf-zone at Del Mar, San Diego County, California, with notes on its morphology. Bulletin of the Southern California Academy of Sciences 102(3):99–107.
- Lea, R. N. 1988. Family Himantolophidae added to the ichthyofauna of the temperate eastern North Pacific. California Fish and Game 74:180–182.
- Love, M. S., and J. K. Passarelli, editors. 2020. Miller and Lea’s Guide to the Coastal Marine Fishes of California. 2nd edition. UCANR Publications, Davis, California, USA.
- Love, M. S., J. J. Bizzarro, A. M. Cornthwaite, B. W. Frable, and K. P. Maslenikov. 2021. Checklist of marine and estuarine fishes from the Alaska–Yukon border, Beaufort Sea, to Cabo San Lucas, Mexico. Zootaxa 5053(1):1–285. https://doi.org/10.11646/zootaxa.5053.1.1
- Luck, D. G., and T. W. Pietsch. 2008. In-situ observations of a deep-sea ceratioid anglerfish of the genus Oneirodes (Lophiiformes: Oneirodidae). Copeia 2008(2):446–451. https://doi.org/10.1643/CE-07-075
- Ludt, W. B., and T. R. Clardy. 2022. First detection of biofluorescence in a deep-sea anglerfish. Journal of Fish Biology 100(3):843–846. https://doi.org/10.1111/jfb.14988
- Lundsten, L., S. B. Johnson, G. M. Cailliet, A. P. DeVogelaere, and D. A. Clague. 2012. Morphological, molecular, and in situ behavioral observations of the rare deep-sea anglerfish Chaunacops coloratus (Garman, 1899), order Lophiiformes, in the eastern North Pacific. Deep Sea Research Part I: Oceanographic Research Papers 68:46–53. https://doi.org/10.1016/j.dsr.2012.05.012
- Pietsch, T. W. 2009. Oceanic Anglerfishes: Extraordinary Diversity in the Deep Sea. University of California Press, Berkley, CA, USA.
- Sabaj, M. H. 2020. Codes for natural history collections in ichthyology and herpetology. Copeia, 108(3):593–669. https://doi.org/10.1643/ASIHCODONS2020
- Stewart, A. L., T. W. Pietsch, J. Moore, and X. Peng. 2023. Upside‐down swimming: in‐situ observations of inverted orientation in Gigantactis, with a new depth record for the Ceratioidei. Journal of Fish Biology 104(3):887–891. https://doi.org/10.1111/jfb.15609
- Tanaka, S. 1918. Figures and descriptions of the fishes of Japan including Riukiu Islands, Bonin Islands, Formosa, Kurile Islands, Korea and southern Sakhalin. Volume 27. Tokyo Printing Company, Tokyo, Japan.