

FULL RESEARCH ARTICLE
Robert E. Pangle1*, Ashehad A. Ali2, and Michael W. Jenkins3
1 MLJ Environmental, 2520 Venture Oaks Way, Suite 310, Sacramento, CA 95833, USA
![]() https://orcid.org/0000-0001-7273-831X
https://orcid.org/0000-0001-7273-831X
2 New Zealand Institute for Bioeconomy Science, AgResearch Group, Lincoln, Canterbury, New Zealand ![]() https://orcid.org/0000-0002-7328-306X
https://orcid.org/0000-0002-7328-306X
3 California Department of Fish and Wildlife, Habitat Conservation Planning Branch, 1010 Riverside Parkway, West Sacramento, CA 95605, USA ![]() https://orcid.org/0009-0000-4718-1230
https://orcid.org/0009-0000-4718-1230
*Corresponding Author: robert.pangle@gmail.com
Published 29 December 2025 • doi.org/10.51492/cfwj.111.18
Abstract
A key aspect related to successful hemp cultivation is an understanding of the physiological performance of hemp and how differing cultivars and genotypes respond physiologically to cultivation practices and environmental stressors. In this study, we examine the leaf-level physiological response of two hemp cultivars (“Boax” and “Cherry-Wine”) across the growing season from late vegetative phase to late flowering, with a specific focus on leaf-level gas exchange, photosynthetic capacity, and cannabinoid content. We observed similar and high photosynthetic capacity for the hemp cultivars in our study during late vegetative and early to mid-flowering phases of crop development. Prior to harvest, we observed a seasonal decline in leaf physiological function and photosynthetic capacity in late flowering for both cultivars, and we conclude that a reduction in photosynthetic capacity along with the onset of plant senescence explains much of the reduced physiological performance that we observed as hemp matured into late flower. Plastic mulching cover over planting rows had no significant effect on leaf physiological function or final cannabidiol (CBD) content, and we attribute this lack of any treatment effect to the fact that plastic mulch cover did not significantly affect canopy microclimate, nor did it significantly affect plant-available soil water in any physiological impactful manner. The Boax and Cherry-Wine cultivars exhibit high physiological performance in relation to photosynthetic capacity and a correspondingly high CBD content; however, despite the inherent productivity of these two cultivars, growers should be cautious when utilizing these hemp cultivars given their potential to exceed tetrahydrocannabinol (THC%) regulatory thresholds during compliance testing.
Key words: cannabinoid potency, hemp, leaf gas exchange, photosynthetic capacity
| Citation: Pangle, R. E., A. A. Ali, and M. W. Jenkins. 2025 Hemp physiological response to seasonal variation in photosynthetic capacity for two high yield cannabidiol (CBD) cultivars. California Fish and Wildlife Journal 111:e18. |
| Editor: Cherilyn Burton, Habitat Conservation Planning Branch |
| Submitted: 26 November 2024; Accepted: 15 January 2025 |
| Copyright: ©2025, Pangle et al. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Funding: Rubisco Research and Consulting, LLC and VIP Farms A NC, LLC provided funding support related to crop establishment and execution of the field experiment. |
| Competing Interests: The authors have not declared any competing interests. |
Introduction
Industrial hemp (Cannabis sativa L.) is an important agricultural crop, which has long been cultivated for fiber and seed (Cherney and Small 2016). Hemp cultivation in the United States has increased dramatically since 2013, following the implementation of hemp cultivation pilot programs at the state level and the passage of the Agricultural Improvement Act of 2018 (i.e., the 2018 Farm Bill), which legalized the production of commercial hemp nationwide at the federal level (Malone and Gomez 2019; Mark et al. 2020). Hemp is legally defined by the 2018 Farm Bill as cannabis (and derivatives of) with ∆9 – tetrahydrocannabinol (THC) levels no higher than 0.3% on a dry weight basis (Mark et al. 2020). In recent years, hemp production has increasingly focused on the cultivation of high yielding cultivars possessing elevated levels of the non-psychoactive cannabis constituent cannabidiol (CBD), and there are many outstanding questions and challenges facing producers in regard to the agronomic best practices required to optimize cultivation success, especially in the highly dynamic CBD oil extract and flower market (Adesina et al. 2020; Aryal et al. 2024). With the recent legalization of industrial hemp in the US, the opportunities for scientific study and research related to the growth and cultivation of hemp have flourished, and researchers are currently examining a myriad of questions related to the optimization of hemp yield and productivity (Backer et al. 2019; Adesina et al. 2020; Sunoj et al. 2023). One key aspect related to successful hemp cultivation is an understanding of the physiological performance of hemp and how differing cultivars and genotypes respond physiologically to specific cultivation practices, edaphic conditions, and environmental stressors that differ by site, climate, and seasonally across crop development (Blandinières and Amaducci 2022; Trancoso et al. 2022). The fixation of carbon dioxide (CO2) into carbohydrates during photosynthesis is one of the primary physiological factors affecting crop growth, productivity, and ultimately harvestable yield (Long et al. 2006; Brestic et al. 2021). Accordingly, the identification and subsequent propagation of genotypes exhibiting high rates of photosynthetic capacity and growth will provide industry and growers with the necessary agronomic data and planting material to enable selection of cultivars having the potential to maximize productivity and yield (Campbell et al. 2019; Blandinières and Amaducci 2022; Trancoso et al. 2022).
Fundamentally, light irradiance, temperature, atmospheric CO2, nutrient availability, and water (H2O) are essential factors affecting photosynthesis (Brestic et al. 2021). Foremost, solar radiation provides the energy that drives the light dependent reactions of photosynthesis, while atmospheric CO2 serves as the substrate and basic building block for carbohydrate synthesis, with the concentration of CO2 in leaf chloroplasts being highly dependent on atmospheric CO2 levels and the diffusion of CO2 into the leaf intercellular spaces via stomatal pores in the leaves (Farquhar and Sharkey 1982; Eichhorn Bilodeau et al. 2019). A consequence of CO2 uptake via stomatal opening is an accompanying loss of H2O to the atmosphere from plant canopies through transpiration—a process which is strongly affected by the vapor pressure deficit (VPD) of the surrounding atmosphere (Grossiord et al. 2020). As such, stomata in leaves serve to regulate the foliar uptake of CO2 for use in carbon fixation by photosynthesis, while also regulating the loss of H2O via transpiration to avoid dehydration and eventual failure of the plant hydraulic system via xylem cavitation and desiccation (Farquhar and Sharkey 1982; Sperry 2000). The ratio of carbon fixation (i.e., net photosynthesis) to transpiration H2O loss at the leaf-level is known as instantaneous H2O use efficiency (WUE), and this metric is often used to identify genotypes and cultivars that have high productivity while also minimizing transpirational H2O losses—a key physiological attribute of genotypes/cultivars that may be more suitable for planting in drier and drought prone sites, or situations where irrigation supplies or infrastructure are limited (Hatfield and Dold 2019; Leakey et al. 2019). Accordingly, measurements of leaf gas exchange parameters provide insightful data for assessing the inherent potential productivity of differing cannabis genotypes and allows for an assessment of how realized plant photosynthesis and productivity varies in response to phenological and environmental change over the course of crop development (Bauerle et al. 2020; Rodriguez-Morrison et al. 2021; Blandinières and Amaducci 2022; Trancoso et al. 2022).
In this study, we examined the leaf-level physiological response and cannabinoid CBD yield of two hemp cultivars across the growing season from late vegetative phase to late flowering and harvest, with a specific focus on leaf gas exchange parameters related to leaf-level CO2 and H2O flux rates (i.e., photosynthetic carbon fixation (leaf-An), stomatal conductance (leaf-gs), transpiration (leaf-EL), and water-use efficiency [WUE]). We also measure and quantify the seasonal change in parameters related to photosynthetic capacity (i.e., the maximum rate for the carboxylation capacity of the enzyme Rubisco (Vcmax), the electron transport rate for the regeneration of RuBP (Jmax), the triose phosphate utilization rate (TPU) in the chloroplast, and light saturated photosynthesis [Amax]). In conjunction with seasonal measurements of hemp physiological performance, we also incorporated an assessment of the leaf-level physiological response and cannabinoid CBD yield of hemp cultivars in response to the common cultivation practice of plastic mulching, which was tested against bare ground cultivation utilizing a randomized block design. Cultivation practices can help optimize cultivation success through influences on plant physiological responses due to modifications and changes in the basic environmental resources and drivers that govern how plants respond to their surrounding environment, and this is especially the case for the fundamental physiological processes related to canopy leaf-gas exchange and crop photosynthesis.
Methods
Study Site and Experimental Design
The study site was located near Robbins in Moore County, NC, USA (35.4114 N, –79.5515 W, 145 m asl.), on well-drained (slope 0–4%), silt loam soils (bulk density =1.11 g cm3). Average annual precipitation ranges from 1,016 to 1,143 mm (Carthage NC, NOAA 2025), with mean monthly high and low temperatures (April to October) averaging 21–32°C and 8–20°C, respectively. We established a randomized block design consisting of six experimental blocks (see Table S1) in July 2019 with a planting of 924 hemp clones (Cannabis sativa L.) using two high CBD yielding (low ∆9-THC) cannabis cultivars “Boax” and “Cherry-Wine”, hereafter referred to as BX and CW respectively, and three differing factors: two levels related to hemp cultivar (i.e., BX vs. CW), two levels of black row bed mulching cover (i.e., plastic mulch cover versus no plastic), and two levels of nutrient fertilizer amendment (see Table S1 and Supplemental Information for more details on the study site and experimental design).
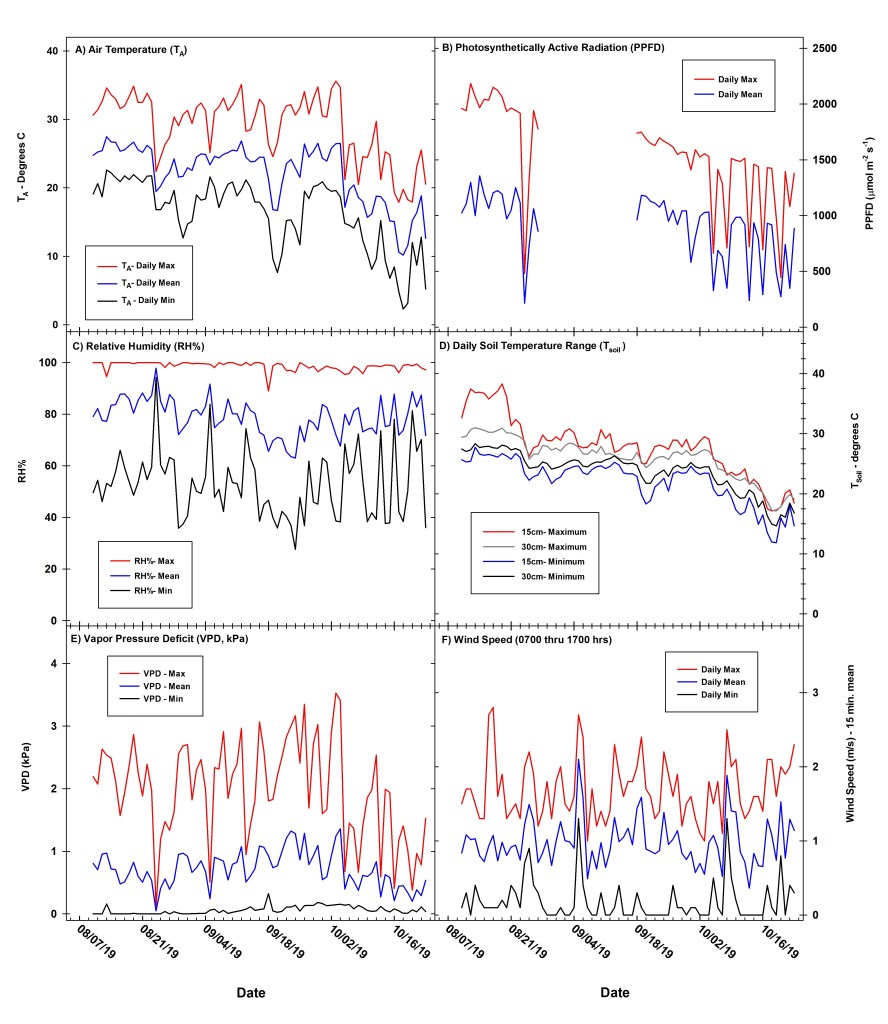
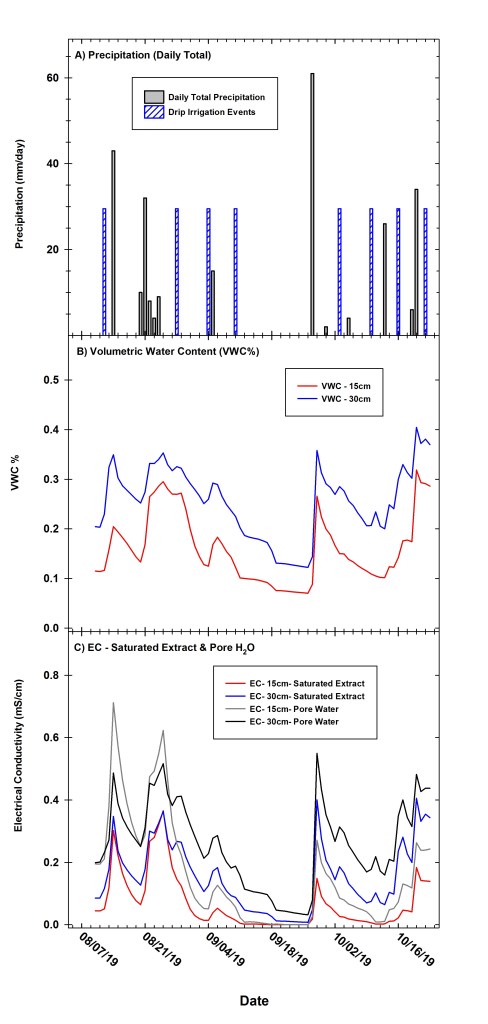
We supplemented natural precipitation with periodic irrigation, and supplemental irrigation events supplied the depth equivalent of 29.2 mm of water into planting beds across all six blocks. Supplemental irrigation via drip tape was applied to the experimental study plot on seven dates during the growing season. In four out of six replicate blocks (blocks #2–5), we injected supplemental irrigation with liquid nutrient fertilizers, whereas the two remaining replicate blocks (blocks #1 and 6) received comparable irrigation levels, but no nutrient additions (again, see Supplemental Information for details). Details related to nutrient amendments and treatments are covered in a separate analysis. We measured environmental conditions during the duration of our study (Figs. 1, 2) using a meteorological station installed within our study site and recorded data at 15-minute intervals including measurements of air temperature (Ta), relative humidity (RH), vapor pressure deficit (VPD), barometric pressure (Patm), photosynthetically active radiation (PAR, also referred to as photosynthetic photon flux density (PPFD), 400–700 nm), wind speed and direction, and precipitation (see Supplemental Information for more details).
Physiological Measurements
We randomly selected 10 plants from each block for repeated physiological measurements over the course of the study, with two plants being selected from each row, which provided a sample size of n = 5 plants for each of the two cultivars within a specific block (Please note: throughout the manuscript, “n” is used to denote the sample size for various measurements or parameters, etc.). In total, this provided n = 60 plants for a repeated time-series of physiological measurements across the study, with treatment factors having the following number of plants in each treatment factor: a) cultivar (n = 60 total, n = 30 “BX” versus n = 30 “CW”), b) plastic mulching cover (n = 40 total, n = 20 “plastic cover” versus n = 20 “no plastic”), split evenly between cultivars, and c) nutrient amendment (n = 40 total, n = 20 “nutrient amendment” versus n = 20 “no amendment”). For each selected plant, we designated an adjacent plant (of the same cultivar and treatment factor) as a back-up to serve as a replacement in case of mortality, disease, or herbivore damage during the study.
To assess changes in leaf-level physiology and physiological responses over the course of our study, we made repeated measures of leaf net photosynthesis (An), leaf stomatal conductance (gs), leaf transpiration (EL), foliar chlorophyll content (CCI), foliar nutrient content (%), leaf temperature (TL), and plant foliar water potential (Y) across a ten week period from 14 August to 17 October 2019 that spanned from the pre-flowering vegetative phase of plant development through flowering to maturation (see Figs. 3, 4). Regarding plant development, flowering initiation was first observed in both cultivars on 24 August (BX) and 25 August (CW), and all 60 plants used in repeated physiological measurements were harvested on 29 October 2019. Harvested biomass included all above ground biomass (AGB, including stems, branches, leaves and inflorescences). We hand-cut plants at ~5cm stem height above ground level, weighed (wet AGB) and hung in an open-air drying barn and dry weights obtained.
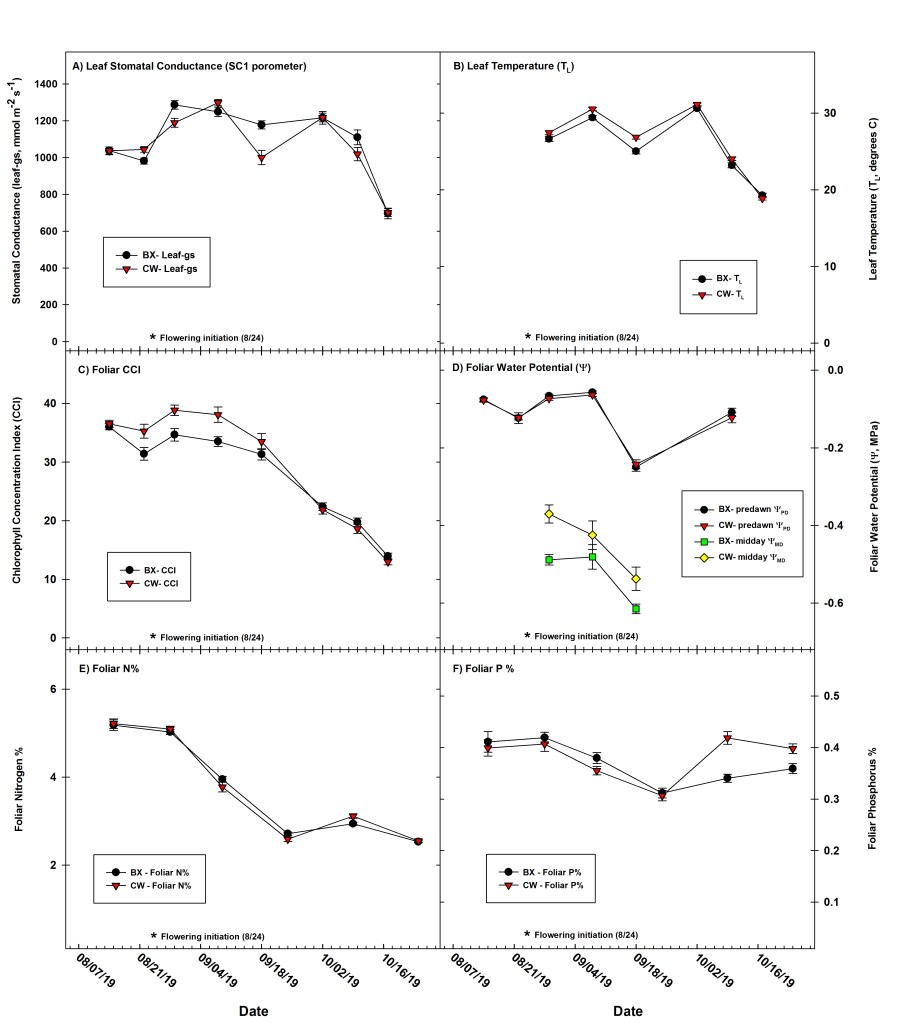

We performed weekly assessments (n = 8, Fig. 3) of leaf-level physiological response across all 60 plants via measurements of leaf-gs(utilizing an SC-1 porometer, METER Group, Inc. USA), foliar CCI (MC-100, Apogee Instruments), and leaf temperature (Fluke, Model 561, infrared thermometer) on fully illuminated leaves in the upper canopy. The SC-1 porometer was calibrated daily before each use, and we made weekly measurements midday between 1100 to 1530 hrs on dry plant canopies under sunny to mostly sunny conditions. For balanced sampling across the experimental design, we measured two plants from each replicate block (one per cultivar) before proceeding to the next block. This pattern was followed across all six blocks for an initial total of n = 12 measurements across all blocks, then the pattern was repeated until all 60 plants were eventually measured. On a subset of weekly measurement days, we assessed plant water status and water stress on n = 16 plants (Fig. 3d) via measurements of predawn water potential (Ypd, n = 6 dates) and midday water potential (Ymd, n = 3 dates) using a Scholander type pressure chamber (PMS Instruments).
We collected foliar tissue samples (upper canopy) for nutrient analysis (≈ 2-week intervals) from late vegetative (15 August) through late flower (24 October) growth phases (n = 24 to 48 plants by date, balanced across cultivar and plastic treatments). We oven-dried samples at 60°C for 48 hrs and shipped to the North Carolina NCDA&CS Agronomy Services Division for laboratory analysis. For potency testing, we collected fresh flower tissue samples from hemp flowers (n = 36 to 48 plants by date, balanced across cultivar and plastic treatments) on three dates corresponding to early-mid flowering (19 September), late flowering (10 October), and senescence/harvest (24 October). Potency analysis was performed by Pinnacle CT, Inc. Testing Services (Westminster, MD) using high-performance liquid chromatography coupled with UV detection. Total CBD and THC% were calculated as follows: total CBD% = (CBD% + CBDA% × 0.877), and total THC% = (THC% + THCA% × 0.877).
In addition to weekly assessments of hemp leaf physiological responses, we performed periodic measurements (Fig. 4) of leaf photosynthesis (An), stomatal conductance (gs), and transpiration (leaf-EL) on fully illuminated leaves in the upper canopy using an LI-6800 portable photosynthesis system equipped with a 3×3 cm clear top leaf chamber (LI-COR Biosciences, Inc.). From the resulting gas exchange data, we also calculated estimates of both instantaneous and intrinsic water use efficiency (WUE) as follows: a) instantaneous WUE = leaf An/EL, and b) intrinsic WUEi =leaf An/gs. Measurements were performed on n = 4 dates (22 August through 15 Oct, both pre and post flower initiation) from late morning through afternoon (1100 to 1600 hrs) on days with sunny to mostly sunny conditions. For each sampling effort utilizing the LI-6800, a total of n = 24 plants were measured (total of n = 96 measurements across the season), with an equal number of plants measured from each cultivar and plastic mulching treatment factor across blocks. Measurements were taken at ambient Ta (ranging from 24 to 36°C depending on date and time of day), ambient RH and VPD (ranging from ~50 to 78%, and ~1.0 to 2.2 kPa respectively), and ambient CO2 concentrations (~400 ppm). Flow rate for the system gas analyzers was set at 500 mmol s-1, and the 3×3 cm clear top leaf chamber light source was set at a saturating PPFD of 1500 to 2000 mmol m-2 s-1, depending on the date when measurements were performed (see Supplemental Information more details on physiological measurements).
Light and CO2 Response Curves
Light response curves (LRC) were measured during late vegetative (19 and 20 August) and mid-late flower (9 October) growth periods. Light response curves were measured using the Light Response Program (with default settings) provided within the Auto Programs function of the LI-6800. Sample size was n = 12 plants for each measurement period, which included plants from each cultivar and plastic mulching treatment. Carbon dioxide response curves were used to measure the response of leaf-An to increasing leaf internal CO2 (Ci) concentrations. This response was measured using the rapidA-Ci response (RACiR) procedure and the AutoLog program function of the LI-6800 (as outlined in Stinziano et al. 2017). Measurement of RACiR curves were performed over the course of three seasonal campaigns including: late vegetative (21–23 August), early flower (10–12 October), and late flower (10 and 15 October). For each measurement campaign, we measured n = 24 plants using the RACiR procedure, including plants from each cultivar and plastic mulching treatment (for a total of n = 72 RACiR measurements over the study duration). See Supplemental Information for a detailed description of the methods and procedures used for both the LRC and RACiR measurements).
Once processed, we used corrected RACiR data (An versus Ci) to obtain Vcmax and Jmax estimates based on the FvCB model (Farquhar et al. 1980) using R open-source software (R version 3.5.3) and the “plantecophys” R package (Ver. 1.4.4, Duursma et al. 2015). The “fitaci” function was used for curve fitting, along with leaf An, TL, PPFD, and Ci (<1000 ppm) as input variables. All parameter outputs were corrected to 25°C during the fitting procedure, and the fit method was set to “bilinear”. All fit parameter values used in the fitting procedure, including a temperature adjusted Michaelis-Menten coefficient for the Farquhar model and photorespiratory CO2 compensation point, are reported in Table 1. Due to the saturation of photosynthesis at high Ci (Long and Bernacchi 2003), the “fit TPU” option in the “fitaci” function was utilized (see Fig. S1 for an example of an A-Ci curve with TPU fitted). Triose phosphate utilization (TPU) was estimated for all A-Ci datasets, and utilizing the TPU fitting option improved overall curve fits at high Ci (P<0.0001 and R2 > 0.94, all fits) and provided for more accurate parameter estimates for Jmax. See Gregory et al. (2021) for a discussion of potential underestimation of Jmax when TPU is not accounted for under situations of photosynthesis saturation at high Ci.
Table 1. Seasonal change in photosynthetic capacity for hemp cultivars at the Robbins experimental field site in 2019, including Vcmax (carboxylation capacity of the enzyme Rubisco), Jmax (electron transport rate for the regeneration of RuBP), TPU (triose phosphate utilization rate), and Amax (light saturated photosynthesis). Data presented are mean values (± 1 SEM).
| Variety | Period | Vcmax (μmol m-2 s-1) | Jmax (μmol m-2 s-1) | TPU (μmol m-2 s-1) | Amax (μmol m-2 s-1) |
| BX | Late Vegetative | 143.2 (2.7) | 325.9 (7.8) | 19.0 (0.31) | 48.8 (1.9) |
| BX | Early Flower | 138.2 (6.7) | 336.3 (19.6) | 19.5 (0.68) | N/A |
| BX | Late Flower | 91.7 (5.3) | 155.3 (7.7) | 9.9 (0.41) | 26.7 (1.5) |
| CW | Late Vegetative | 136.2 (2.6) | 323.4 (8.3) | 19.3 (0.27) | 47.6 (1.3) |
| CW | Early Flower | 135.2 (4.9) | 338.7 (15.8) | 19.4 (0.45) | N/A |
| CW | Late Flower | 95.4 (6.2) | 161.1 (10.5) | 10.0 (0.47) | 27.3 (3.2) |
Statistical Analysis
We analyzed physiological data using GraphPad Prism version 9.2.0 (GraphPad Software, LLC). Statistical tests for significance between physiological variables and seasonal/treatment effects were performed using a Mixed Effects Model (REML) with a repeated measures analysis structure. In each mixed effects model, the dependent variable was tested for significance against the fixed effects related to Date, Cultivar, and Plastic Mulching treatment. For the repeated measures portion of the analysis, date was the repeated factor and plant ID was treated as the subject in the random effects. Main and second order interaction effects were tested for significance at the P = 0.05 threshold. In the event of significant 2nd order interactions between fixed effects, non-significant treatment factors were pooled and a two-way ANOVA with repeated measures and multiple comparisons (by date) was used to identify the specific 2nd order interaction that was present. A Bonferroni post hoc correction was used in all instances where multiple comparisons were performed. Results are reported as the mean value plus-or-minus one standard error of the mean (SEM). The two experimental factors explicitly examined in our analysis are those related to cultivar and the effect of plastic mulching treatment. Plastic mulching was observed to be a non-significant first order treatment factor for all parameters assessed except for Ymd, and statistically significant second order interactions related to plastic mulching were infrequent or were physiologically insignificant when they occurred. Accordingly, data results were subsequently pooled and presented by date and cultivar only (i.e., the plastic mulching factor was removed from the data presentation).
Results
Environmental Variables
Seasonal trends in environmental variables across the study period are presented in Figures 1 and 2. Daily mean, maximum and minimum Ta averaged 22.1, 29.0, and 16.2°C, respectively, from early August through late October 2019 at the study site (Fig. 1a). Maximum daily PPFD averaged 2000 to 2200 mmol m-2 s-1 in mid-August, decreasing to a daily maximum of 1400 to 1500 mmol m-2 s-1 by mid-October 2019 (Fig. 1b). Daily mean RH averaged 78.8 % across the study period (Fig. 1c), while daily maximum and minimum VPD averaged 2.0 and 0.061 kPa (Fig. 1e). Daily maximum and minimum Tsoil (treatments pooled) averaged 27.9 and 22.1°C at 15cm depth, and 26.0 and 23.7°C at 30 cm, respectively (Fig. 1d). Precipitation measurements began on 10 August with a total of 280 mm of rainfall received through late October 2019 (Fig. 2a). We supplemented natural precipitation with a total of eight irrigation events that supplied the depth equivalent of approximately 29.2 mm of water per event (Fig. 2a) with changes in soil VWC (m3 m-3) at varying depths closely tracking natural precipitation events (Fig. 2b). Across the duration of the study, soil VWC averaged 15.9% and 25.4% at 15 and 30 cm depth, respectively (Fig. 2b). Our study site experienced a notable dry-down period between 6 September and 26 Sept, a 3-week period when no precipitation occurred and no supplemental irrigation was applied (Fig. 2b). During this dry-down period, average soil VWC at 15 and 30 cm depth declined to 7.0% and 12.3%, respectively. Finally, soil electrical conductivity (EC) exhibited a strong relationship with changes in soil VWC (Fig. 2b,c), with ECe and ECw at varying depths increasing to maximum values of 0.4 to 0.7 mS cm-1,respectively,post precipitation (Fig. 2c).
Leaf Gas Exchange – An, gs, EL, and WUE
Seasonal patterns of An, gs, and EL measured with the LI-6800 (Fig. 4) followed a similar trend to leaf gas exchange (leaf – gs) measured with the SC-1 porometer (Fig. 3a). Maximum rates of gas exchange were observed during the late vegetative and early to mid-flowering portion of plant development, with gas exchange rates declining as late summer transitioned into fall (Figs. 3, 4). Leaf An, gs, and EL (measured with LI-6800) were all found to significantly vary by date (P < 0.0001, Fig. 4). However, cultivar type (P = 0.90, P = 0.53, and P = 0.30, respectively for An, gs, and EL) and plastic mulching treatment (P = 0.11, P = 0.51, and P = 0.98 respectively for An, gs, and EL) were not found to be significant, and no significant 2nd order interactions were observed for any of the three aforementioned gas exchange variables measured using the LI-6800 (Fig. 4). Similarly, weekly leaf-gs (measured via the SC-1 porometer, Fig. 3a) varied significantly by date (P < 0.0001), but cultivar type (P = 0.15) and plastic mulching treatment (P = 0.08) were also not found to be significant treatment factors, and although a significant 2nd order interaction between date × cultivar (P < 0.0001) was observed for the 18 September measurements (Fig. 3a), overall mean leaf-gs still averaged close to 1.0 mol m-2 s-1 in all instances.
Over the course of the study, leaf-An ranged from a mean high of 42.4 (SEM = 1.60) mmol m-2 s-1 for BX cultivar plants on 11 September to a mean low of 21.9 (1.54) mmol m-2 s-1 for the BX grouping of plants on 15 October (Fig. 4a). Leaf gs and EL (LI-6800) ranged from a mean high of 1.15 (0.033) and 0.016 (0.0007) mol m-2 s-1 respectively for plants in the BX cultivar group on 22 August to a mean low of 0.504 (0.056) and 0.0069 (0.0007) mol m-2 s-1 respectively for the CW cultivar grouping of plants on 15 October (Fig. 4b,c). Instantaneous water use efficiency (i.e., WUE, leaf An/EL) generally ranged from a mean of 2.6 to 3.5 mmol CO2/mmol H2O by cultivar across the season (Fig. 4d). Instantaneous water use efficiency varied significantly by date (P = 0.02), but cultivar type and plastic mulching treatment were not found to be significant treatment factors (P > 0.25, all instances), and no significant 2nd order interactions were observed.
Photosynthetic Capacity – Amax, CCI, Vcmax, Jmax, and TPU
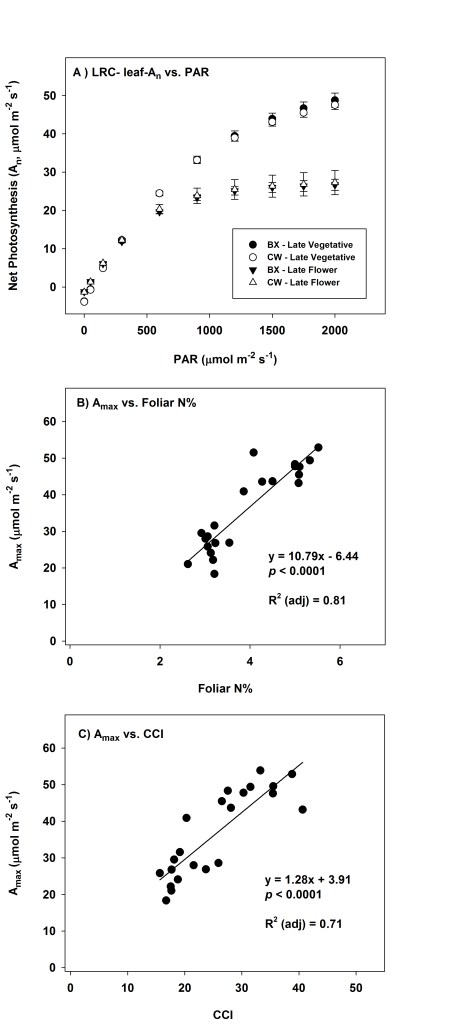
Light saturated photosynthesis (Amax) averaged 48.8 (SEM = 1.9) and 47.6 (1.3) mmol m-2 s-1 respectively for BX and CW cultivars measured during late vegetative growth in mid-August 2019 at ambient CO2 and a saturating PPFD of 2,000 µmol m-2 s-1 (Fig. 5a, Table 1). A seasonal decline in photosynthetic capacity was observed, with Amax declining to 26.7 (1.5) and 27.3 (3.2) mmol m-2 s-1 respectively for BX and CW cultivars measured during late flowering in mid-October 2019 at ambient CO2 and saturating PPFD of 2,000 µmol m-2 s-1 (Fig. 5a, Table 1). Date was a significant factor (P < 0.0001) in regard to seasonal declines in Amax, whereas cultivar and plastic mulching were not significant factors (P = 0.77 and P = 0.27, respectively). No significant 2nd order interactions were noted in regard to Amax. Seasonally, foliar chlorophyll content index (CCI, unit-less) also exhibited a similar decline from a mean high of 38.8 (SEM = 0.87) for the CW cultivar plants on 29 August to an average low of 13.0 (0.51) for the CW cultivar plants on 17 October (Fig. 3c). Chlorophyll content index was found to significantly vary depending on date (P < 0.0001), but cultivar type (P = 0.08) and plastic mulching treatment (P = 0.07) were not found to be significant treatment factors. Likewise, foliar N% followed a similar seasonal pattern as Amax and CCI, varying from 5.2% (SEM = 0.08) across all treatments on 15 August to an average low of 2.5% (0.021) across treatments on 24 Oct, just prior to harvest (Fig. 3e). Foliar N% and P% (Fig. 3e, 3f) varied significantly by date (P < 0.0001 both instances), but cultivar type (P = 0.60 and P = 0.06, respectively) and plastic mulching (P= 0.88 and P= 0.78, respectively) were not found to be significant treatment factors. A significant and strong relationship (P < 0.0001) between Amax and both foliar N% (R2 = 0.81, Fig. 5b) and CCI (R2 = 0.71, Fig. 5c) was observed seasonally. While N% and CCI were also observed to have a significant and positive relationship across seasonal measurements (P < 0.0001, Fig. S2:panels e and f), the potential use of CCI as a simple predictor of leaf N% was deemed not to be practical given the weak linear relationship between the two variables (R2 = 0.40).
Regarding Vcmax, Jmax and TPU, measurement date was observed to be significant (P < 0.0001) in all instances. Cultivar type was not a significant treatment factor in regard to Vcmax, Jmax and TPU (P = 0.77, P = 0.94, and P = 0.51 respectively), and neither was plastic mulching (P = 0.80, P = 0.77, and P = 0.80, respectively), and no significant 2nd order interactions were observed regarding Vcmax, Jmax and TPU. The maximum carboxylation capacity of Rubisco (Vcmax) averaged 139.7 (SEM = 1.9) and 136.7 (4.0) mmol m-2 s-1 during mid-August (i.e., late vegetative) and mid-September (i.e., early flower) ACi curve measurement campaigns (Table 1, cultivars pooled), while the maximum rate of electron transport (Jmax) averaged 324.7 (5.6) and 337.5 (12.3) mmol m-2 s-1 over this same time frame, respectively. Triose phosphate utilization (TPU) averaged 19.2 (0.21) and 19.5 (0.39) mmol m-2 s-1 during late vegetative and early flower ACi curve measurement campaigns. By late flowering, Vcmax, Jmax, and TPU all declined to an overall mean of 93.6 (4.0), 158.2 (6.4), and 10.0 (0.30), respectively, by mid-October (Table 1, cultivars pooled). Both Vcmax and Jmax exhibited a significant and positive relationship with N% and CCI (P < 0.0001, Fig. S2: panels a–d); however, the strength of the linear relationships was less robust in some instances (R2 ranged from 0.43 to 0.60).
Foliar Temperature (TL) and Water Status (Y)
Leaf TL (Fig. 3b) was found to significantly vary by date (P < 0.0001) and cultivar type (P = 0.02), but plastic mulching treatment was an insignificant factor (P = 0.49). Significant 2nd order interactions for TL were observed between date × cultivar (P = 0.0004) and date × plastic mulching (P = 0.02) treatment factors. Post-hoc analysis revealed a significant difference (P < 0.05) in TL between the plastic and no-plastic mulching treatments on 18 Sept; however, the average TL difference between the two groups was only 0.87°C and was subsequently considered insignificant from a physiological perspective. Daily mean TL varied with changes in Ta, ranging from a mean high of 31.1 (SEM = 0.16)°C for CW cultivar plants on 2 October to a daily mean low of 18.9 (0.22)°C for CW plants on 17 October (Fig. 3b).
Foliar Ypd was found to significantly vary by date (P < 0.0001), but cultivar type (P = 0.44) and plastic mulching treatment (P = 0.78) were not found to be significant treatment factors, and no significant 2nd order interactions were observed. Predawn foliar water potential (Ypd) declined during the late summer dry-down, decreasing from a high of -0.057 (SEM = 0.003) MPa for BX cultivar plants on 7 September to an average low of -0.25 (0.01) MPa for BX plants on 17 September (Fig. 3d). Midday foliar water potential (Ymd) also declined with soil moisture during dry-down, decreasing from a high of -0.37 (0.023) MPa for CW cultivar plants on 29 August to an average low of –0.62 (0.012) MPa for BX cultivar plants on 18 September (Fig. 3d). Foliar Ymd was found to significantly vary by date (P < 0.0001), cultivar (P = 0.01), and plastic mulching treatment (P = 0.03), and a significant 2nd order interaction (P = 0.006) was observed between date × plastic mulching treatment factors. Across all observations, the BX cultivars averaged a Ymd of –0.53 (0.013) MPa compared to CW cultivars which averaged –0.44 (0.015) MPa.
Potency and Yield
Prior to harvest at 61 days post-flowering initiation, total CBD and THC% for hemp flower tissue averaged 19.6 % (SEM = 0.67) and 0.80 % (0.02) for BX cultivars and 15.3 % (0.39) and 0.57 % (0.02) for CW cultivars respectively (Table 2). Total CBD and THC % exhibited a decline relative to concentrations observed at 47 days post flowering initiation, and CBD and THC concentrations were significantly higher for BX compared to CW cultivars (P < 0.0001, both instances) during both the late flowering and senescence/harvest sampling periods (Table 2). Plastic mulching treatment did not have a significant effect on total CBD and THC concentrations for either cultivar during late flowering and harvest sampling (P = 0.48 and 0.91, respectively), nor were there any significant 2nd order interactions between plastic mulching and cultivar in regard to total CBD and THC%. Total CBD % was regressed against leaf-An and foliar CCI to determine if a significant relationship existed between growing season leaf-level physiological factors and total CBD production; however, no relationship was observed as regression comparisons across differing timeframes and averaging periods were all found to be non-significant (R2 values ranged from 0.0005 to 0.053). Finally, regarding biomass, total aboveground harvestable biomass (AGB, i.e., stems, branches, leaves and inflorescences) averaged 1.12 (0.05) and 2.15 (0.13) kg plant-1 for BX and CW cultivars respectively (Table 2). There was a significant difference between cultivars in regard to AGB (P < 0.0001), but plastic mulching had no significant effect (P = 0.43) and there were no significant 2ndorder interactions present between plastic mulching treatments and cultivars.
Table 2. Flower potency and harvest yield for Boax (BX) and Cherry-Wine (CW) hemp cultivars at the Robbins experimental field site in 2019. Totala CBD and THC% potency for hemp flower sampled across n = 3 dates from early-mid flowering (19 September) to late flowering (10 October) and senescence (24 October). Harvested biomass (kg/plant, 29 October) reflects total aboveground biomass (AGB) dry weight including stem, branch, leaf, and inflorescence. Data presented are mean values (± 1 SEM).
| Variety | Flowering stage | Days post flower initiation | Flower- totala CBD (%) | Flower- total1 THC (%) | Harvest AGB (kg/plant) |
| BX | Early-Mid Flower | 26 | 11.5 (0.31) | 0.33 (0.02) | N/A |
| BX | Late Flower | 47 | 20.8 (0.60) | 0.92 (0.03) | N/A |
| BX | Senescence & Harvest | 61 | 19.6 (0.67) | 0.80 (0.02) | 1.12 (0.05) |
| CW | Early-Mid Flower | 26 | 7.6 (0.53) | 0.16 (0.03) | N/A |
| CW | Late Flower | 47 | 17.0 (0.63) | 0.70 (0.02) | N/A |
| CW | Senescence & Harvest | 61 | 15.3 (0.39) | 0.57 (0.02) | 2.15 (0.13) |
Discussion
Physiological Performance of Cultivars
The hemp cultivars in our study exhibited similar and high photosynthetic capacity (Amax) both prior to and following flower initiation (Figs. 4, 5, Table 1), with average An rates upwards of 48 mmol m-2 s-1 observed for plants growing at ambient atmospheric CO2 concentrations and saturating light in an outdoor field setting. Previous studies on indoor/greenhouse grown cannabis have documented the high An of cannabis cultivated under both ambient and elevated/enriched CO2 air environments (Bazzaz et al. 1975; Lydon et al. 1987; Chandra et al. 2008; Jenkins and Livesay 2021; Rodriguez-Morrison et al. 2021). Likewise, industrial hemp has been documented to possess a high An (Tang et al. 2017; Tang et al. 2018; Bauerle et al. 2020), especially under conditions of high foliar nitrogen (N) content and no water stress. Notably, for hemp grown under ambient CO2 concentrations in an outdoor agricultural setting, LiCor Biosciences (2022) reported quite similar An rates to those observed in our study at comparable leaf internal CO2 concentrations (Ci). The high Amax of cannabis necessitates a correspondingly high degree of stomatal opening and leaf-gs to provide for adequate diffusion of CO2 into the leaf intercellular spaces during periods of high photosynthetic carbon fixation (Wong et al. 1979; Reich et al. 1999; Gilbert et al. 2011), and this particular correlation in leaf physiological traits is especially critical for C3 plants growing at ambient atmospheric CO2 concentrations in outdoor agronomic settings. Observed leaf-gs and EL rates in our study were similar between cultivars and notably higher than rates reported by Chandra et al. (2008, 2015), Lydon et al. (1987), Murovec et al. (2022), and Shiponi and Bernstein (2021), but measured leaf-gs in our study were similar to maximum leaf-gs reported by Jenkins (2021) and LiCor Biosciences (2022).
Instantaneous and intrinsic WUE in our study did not differ significantly between cultivars, with instantaneous WUE generally averaging between 2.6 to 3.5 mmol CO2 /mmol H2O (Fig. 4d), slightly lower than many WUE observations reported by Chandra et al. (2008, 2015), Murovec et al. (2022), and Tang et al. (2018) for non-drought stressed cannabis plants. The lower WUE we observed compared to these aforementioned studies can partially be attributed to high rates of leaf-gs and a subsequently high degree of transpirational water loss which occurred in our study (Fig. 4b,c), despite the high rates of leaf-An that were measured concurrently (Fig. 4a). Water stressed plants often exhibit an increase in WUE, and this response was reported by Tang et al. (2018) in drought stressed cannabis where WUE increased from 4.0 in well-watered plants up to 7.53 mmol CO2 /mmol H2O in plants exposed to long term water shortage; however, the observed increase in WUE came at a high cost to productivity, as canopy gross photosynthesis declined by 68% in droughted plants compared to well-watered cannabis plants in that particular study (Tang et al. 2018). We only experienced one short time period (mid-September 2019) where 15cm and 30cm soil VWC declined below 10% and 20% respectively, and plants began to show a slight decline in both YPD and YMD (Figs. 2b and 3d). While plants did exhibit a slight decline in both YPD and YMD during this temporary soil dry-down (down to approx. -0.6 MPa), measured Y (Fig. 3d) in our study was much higher than values reported by Caplan et al. (2019) for experimentally drought stressed cannabis where foliar Y ranged from -1.0 to -1.5 MPa and drought stressed cannabis plants experienced a 30 to 40% decline in leaf photosynthesis compared to non-droughted controls. Additionally, our weekly to bi-weekly measurements of leaf-gs (Fig 3a) further indicate that plants did not experience significant or prolonged water stress over the course of our study, as plants maintained quite high rates of leaf-gs (e.g., > 900 mmol m-2 s-1) well into flowering stage, only declining prior to harvest in late flowering (Figs. 3a and 4b). Consequently, we surmise that water stress was not a significant constraint on leaf gas exchange for either cultivar during our study. The possibility that the two cultivars in our study may have differed in their sensitivity to water stress is an intriguing question, but we did not experience severe enough declines in plant available water (especially at depth) to detect and assess that question in our study. In future comparative studies of this type, the addition of a drought stress factor in the experimental design framework would provide the ability to fully elucidate the drought sensitivity and WUE of differing cultivars.
Seasonal Variation in Photosynthetic Capacity
Seasonal declines in An and Amax were observed in our study as plants matured towards the late flowering stage (Figs. 4a and 5), and these declines in leaf-level productivity were accompanied by declines in Vcmax, Jmax, foliar N%, and CCI over this same period (Fig. 3, Table 1, Fig. S2). We did not detect significant differences in leaf gas exchange or photosynthetic capacity between the late vegetative and early flowering periods of crop growth (Figs. 3a and 4, Table 1), with initial declines in both N% and CCI detected approximately two- and three-weeks post flower initiation respectively for both cultivars (Fig. 3c,e). Cannabis has been documented to respond positively to high levels of incident light, with leaf-An rates increasing with PPFD levels up to 1,500 to 2,000 umol m-2 s-1, especially in specific genotypes/cultivars possessing high foliar N content (Chandra et al. 2015; Tang et al. 2017; Jenkins and Livesay 2021). While positive responses to increasingly high levels of incident light occur in cannabis, photosynthesis can also reach a light saturation point (LSP), wherein the photosynthetic response plateaus, and no additional photosynthetic response occurs as PPFD increases (Farquhar et al. 1980; Long et al. 2006). In relation to cannabis, seasonal variation in Amaxand LSP has been documented in studies by Bauerle et al. (2020) and Rodriguez-Morrison et al. (2021), where Amax and LSP declined in the latter stages of plant growth and flowering. We observed similar physiological changes in our study in relation to seasonal changes in LSP and Amax, with both cultivars exhibiting increasing An in late vegetative/early flowering up to PPFD of 2,000 mmol m-2 s-1, then a decline in photosynthetic capacity in late flower as leaf An reached LSP at a PPFD range of 1250 to 1500 mmol m-2 s-1 (Fig. 5a). While seasonal declines in temperature, foliar chlorophyll/CCI, foliar N content, and PPFD occurred concurrently as plants matured into late flowering at our site, it is the decline in photosynthetic capacity as evidenced by decreases in Vcmax, Jmax, and subsequently Amax that is particularly notable (Table 1). Subsequently, we posit that this reduction in photosynthetic capacity, in conjunction with the physiological age of foliar tissues and the potential onset of plant senescence, explains much of the reduced physiological performance we observed as hemp matured into late flower (Rodriguez-Morrison et al. 2021). Similar patterns were observed by Bauerle et al. (2020), where decreases in photosynthetic capacity were observed for cannabis leaves over a 42-day period following full leaf expansion, with a progressive decline in Amax, Vcmax, and Jmax observed post leaf expansion regardless of crown position or light intensity. With regard to photosynthetic capacity, crop species have been documented to possess high Amax, Vcmax, and Jmax values (see Tang et al. 2017; Miner and Bauerle 2019; Saathoff and Welles 2021), and the hemp cultivars in our study exhibited a similar high photosynthetic capacity as evidenced by high Vcmax and Jmax parameter estimates and correspondingly high Amax at ambient CO2 concentrations and high PPFD. Triose phosphate utilization estimates in our study ranged from the mid- to higher end of many reported values in the literature (Wullschleger 1993; Kumarathunge et al. 2019), and for studies examining photosynthetic capacity in hemp (Tang et al. 2017; Bauerle et al. 2020). Notably, there is an ongoing debate in the modeling community regarding the importance of incorporating TPU (and/or TPU-limitation) into terrestrial biosphere models, with some authors suggesting limitations may be observed under conditions of low temperature, high light, and high CO2 (Lombardozzi et al. 2017), while others argue that TPU limitations are mostly negligible at current atmospheric CO2 concentrations (see Kumarathunge et al. 2019; Rogers et al. 2021).
Seasonal declines in Amax, Vcmax, and Jmax have been observed to correlate with declines in foliar N% in sunflower and maize (Miner and Bauerle 2019), and Rodriguez-Morrison et al. (2021) postulated that a decline in chlorophyll content over the course of crop development likely contributed to observed reductions in hemp photosynthetic capacity in their study. Foliar N content is well correlated with Amax in many plants, as foliar N is a critical component in many proteins and enzymes related to carbon metabolism and carboxylation capacity in the Calvin cycle, and a significant proportion of foliar N is also located in the chloroplasts and thylakoid membranes which are responsible for light energy capture during photosynthesis (Evans 1989; Chandra et al. 2011; Peterson et al. 1999). Accordingly, total leaf N has been shown to be well correlated in many instances with leaf chlorophyll content, Ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco) enzyme levels, and electron transport capacity related to RuBP regeneration (Evans 1989; Walker et al. 2014). As observed in previous studies (Miner and Bauerle 2019; Rodriguez-Morrison et al. 2021), we noted significant declines in foliar CCI and N% (Fig. 3c,e), and subsequently photosynthetic capacity as hemp plants transitioned into late flowering and approached harvest/senescence (Fig. 5b,c, Table 1, Fig. S2).
Fertilization studies of cannabis (Malceva et al. 2011; Tang et al. 2017; Tang et al. 2018) assessing the effects of N fertilization on photosynthetic capacity and performance have reported significant increases in leaf N content, chlorophyll content, and photosynthetic response to increased N fertilization, with Malceva et al. (2011) observing leaf N content values that ranged upwards of 4–5% in high N treatments, values comparable to foliar N levels observed in hemp plants in the late vegetative and early flowering phases of our study (Fig. 3e). Similarly, Tang et al. (2017) reported significant increases in measured and modeled photosynthetic capacity (upwards of 30–40 mmol m-2 s-1) at higher specific leaf nitrogen (SLN) levels of 1.5–2.5 g N m-2 for plants growing at high light intensity (i.e., PPFD = 2000 mmol m-2 s-1), values comparable to SLN estimates in our study during late vegetative and early flowering periods (SLN approximations ranged from 1.7 to 2.9 g N m-2 in our study). We should note that SLN was not measured directly in our study but estimated based on observed leaf N content (our study) and an assumed specific leaf weight (SLW) of ~45–55 g m2 based on published cannabis values in the literature (see Rodriguez-Morrison et al. 2021 and Moher et al. 2022 for cannabis SLW values). Given the comparably high level of leaf N and associated chlorophyll content (CCI) observed early in our study (Fig. 3c,e), we conclude that soil N availability and subsequently foliar N were not limiting (Cockson et al. 2019; Saloner and Bernstein 2020), and more than adequate for plants to exhibit the high photosynthetic capacity that we observed under conditions of high light intensity, optimal temperature, and low water stress during the late vegetative and early flowering phase of development. Likewise, foliar P content was considered not to be a limiting factor in regard to photosynthetic capacity and growth, as mean foliar P% was > 0.3% in most all instances and periods (Fig. 3f), above the threshold for which P limitation has been documented to occur in cannabis (Cockson et al. 2020; Shiponi and Bernstein 2021; Veazie et al. 2021).
Photosynthesis is also heavily influenced by temperature, as rates of many biochemical and enzymatic functions during carbon fixation are highly temperature dependent, with many species having photosynthetic optima in regard to temperature, beyond which the effects of temperature dependent processes such as photorespiration and increased cellular respiration serve to limit An, especially for C3 crop species (Long et al. 2006; Moore et al. 2021). In our study, measurements of An in the late vegetative and early flowering stages of plant development (i.e., August and September) were performed under well-watered conditions and high PPFD (Figs. 1b and 2), with maximum daily Ta and TL that averaged in the 30 to 34°C range (Figs. 1a and 3b). Previous work by Chandra et al. (2008, 2011) and Tang et al. (2017) report an optimum temperature for An in cannabis to occur in the 25–35°C range for plants measured under high PPFD (1500–2000 mmol m-2 s-1) and ambient CO2 conditions (350–400 ppm). Notably, modeling simulations performed by Tang et al. (2017) predict An rates upwards of 40 mmol m-2 s-1 for hemp plants grown under ambient CO2 conditions and canopy temperatures in the 30–35°C range, rates comparable to those observed in our study under similar Ta, PPFD, and foliar N levels. Tang et al. (2017) also modeled Vcmax, Jmax, and Tp(i.e., TPU) for hemp based on the photosynthesis model of Farquhar et al. (1980), and reported steadily increasing Vcmax and Tprates across a Ta range from 10 to 40°C, whereas Jmax for hemp plateaued across a Ta range of 30 to 40°C. Accordingly, we infer that ambient temperatures at our study site during August/September measurements were within an optimal range for assessing hemp physiological performance in an outdoor agronomic setting, especially in relation to measuring the photosynthetic potential for hemp plants under conditions of high light intensity, low water stress, and high N availability. Hemp leaf-An rates at our site decreased in late flowering as plants approached full maturity and harvest (i.e., October), with average leaf-An rates declining into the 20–30 mmol m-2 s-1 range as maximum Ta and TL declined into the 18–25°C range, with observed leaf-An declines matching well with predicted hemp photosynthesis responses in the model simulation performed by Tang et al. (2017) for similar temperature declines
THC Potency and USDA Guidelines
It should be noted that during flowering and maturation, both BX and CW flower tissue exceeded the 0.3% concentration limit for THC% on a dry weight basis, with BX exceeding the threshold only 26 days post flower initiation (Table 2). Thus, both cultivars in our study surpassed the THC% threshold for hemp classification as outlined by the USDA and the 2018 Farm Bill (Mark et al. 2020). Similar compliance concerns have been observed for industrial hemp in other studies, with Linder et al. (2022) reporting total THC% levels exceeding the 0.3% threshold within ~40 days post flower initiation for the Cherry-Wine cultivars in their study. Notably, the Boax cultivar has been classified as a prohibited variety by some U.S. states, with, for example, Pennsylvania prohibiting the Boax hemp variety after repeated instances where it tested above 0.3% THC and another instance where it tested >1.0% in regard to compliance testing (Pennsylvania Department of Agriculture 2021). In this instance, Pennsylvania did not prohibit the hemp variety Cherry-Wine (as of 2022), but both Pennsylvania and Kentucky have listed it as a variety of concern after repeated instances of potency testing above the 0.3% THC threshold. Thus, while both cultivars in our study exhibited quite high photosynthetic capacity and productivity potential, growers should be cautious when utilizing these two particular hemp cultivars given their potential to exceed THC% allowance thresholds during compliance testing by state-level agencies (Suchoff et al. 2022).
While AGB for CW cultivars exceeded harvested yield for BX cultivars by a large margin in our study (Table 2), it should be noted we utilized total dry AGB to assess yield, and relative yield differences between the two cultivars could differ from our current results if inflorescence biomass alone had been used to assess cultivar yield. For example, harvested AGB in our study exceeded biomass values reported by Linder et al. (2022) for BaOx and Cherry-Wine hemp varieties, but aboveground biomass in the Linder et al. (2022) study reflected inflorescences and leaves only, which helps explain why they report much lower biomass yields compared to our study at a similar harvest schedule (i.e., days post flower initiation). Likewise, Linder et al. (2022) reported lower CBD and THC% compared to concentrations observed in our study, and this may reflect the fact that cannabinoid testing in our study was performed on flower tissue samples alone, as opposed to testing of pooled inflorescence and foliar tissue in the Linder et al. study. The concentration of CBD at harvest is a critical factor to consider when interpreting yield results, and while total AGB yield was lower for BX cultivars in our study, this was partially offset by a higher CBD % in BX relative to CW cultivars (Table 2). Concentrations of CBD for both cultivars in our study were observed to be at the high end of reported values for various regionally grown hemp varieties (Chen et al. 2021; Chiluwal et al. 2023; Suchoff et al. 2022).
Summary
We observed a high photosynthetic capacity (i.e., Vcmax, Jmax, and Amax) for the hemp plants examined in our study, along with observations of correspondingly high rates of leaf-An, gs, and EL during late vegetative and early to mid-flowering phases of crop development, a period when high PPFD, optimal temperatures, high foliar nitrogen content, and lack of plant water stress provided favorable conditions for high photosynthetic carbon gain. There were no significant differences in leaf gas-exchange rates between cultivars in our study, with both cultivars exhibiting very similar levels of leaf physiological function to each other throughout the study. We observed a decline in leaf physiological function and photosynthetic capacity in late flowering for both cultivars leading up to harvest, and we conclude that a reduction in photosynthetic capacity in conjunction with the potential onset of plant senescence explained much of the reduced physiological performance that we observed as hemp matured into late flower at our study site. Plastic mulching had no significant effect on leaf physiological function or final CBD content during the course of our study, and we attribute this lack of any treatment effect to the fact that plastic did not significantly alter the canopy microclimate for hemp in our study, nor did it significantly affect plant-available soil water in any physiologically impactful or consequential manner. Given that both Boax and Cherry-Wine exhibit high physiological performance in relation to photosynthetic capacity and a correspondingly high CBD content, we conclude that both cultivars provide growers with superior planting options in their efforts to maximize productivity and economic returns for hemp grown under similar environmental and site conditions. However, despite the inherent productivity of these two cultivars, growers should be cautious when utilizing such hemp cultivars, given their potential to exceed THC% allowance thresholds during regulatory compliance testing.
Acknowledgements
We thank Rubisco Research and Consulting, LLC and VIP Farms A NC, LLC for funding support related to crop establishment and execution of the field experiment, and we thank Elijah Henry and Farlen Mischel for field assistance with data collection and crop cultivation efforts. Data processing, analysis, and subsequent writing of the manuscript were not funded by any entity or grant.
Declaration of Generative AI and AI- assisted technology in the writing process
The authors did not use Generative AI or AI-assisted technology in the writing of this manuscript.
Literature Cited
- Adesina, I., A. Bhowmik, H. Sharma, and A. Shahbazi. 2020. A review on the current state of knowledge of growing conditions, agronomic soil health practices and utilities of hemp in the United States. Agriculture 10(4):129.
- Aryal, K., T. Maraseni, T. Kretzschmar, D. Chang, M. Naebe, L. Neary, and G. Ash. 2024. Knowledge mapping for a secure and sustainable hemp industry: a systematic literature review. Case Studies in Chemical and Environmental Engineering 9:100550.
- Backer, R., T. Schwinghamer, P. Rosenbaum, V. McCarty, S. Eichhorn Bilodeau, D. Lyu, M. B. Ahmed, G. Robinson, M. Lefsrud, O. Wilkins, and D. L. Smith. 2019. Closing the yield gap for cannabis: a meta-analysis of factors determining cannabis yield. Frontiers in Plant Science 10:495. https://doi.org/10.3389/fpls.2019.00495
- Bauerle, W. L., C. McCullough, M. Iversen, and M. Hazlett. 2020. Leaf age and position effects on quantum yield and photosynthetic capacity in hemp crowns. Plants 9(2):271.
- Bazzaz, F. A., D. Dusek, D. S. Seigler, and A. W. Haney. 1975. Photosynthesis and cannabinoid content of temperate and tropical populations of Cannabis sativa. Biochemical Systematics and Ecology 3(1):15–18.
- Beck, H., N. Zimmermann, T. McVicar, N. Vergopolan, A. Berg, and E. F. Wood. 2018. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Scientific Data 5:180214.
- Blandinières, H., and S. Amaducci. 2022. Agronomy and ecophysiology of hemp cultivation. Pages 89–125 in D. C. Agrawal, R. Kumar, and M. Dhanasekaran, editors. Cannabis/Hemp for Sustainable Agriculture and Materials. Springer Singapore, Singapore.
- Brestic, M., X. Yang, X. Li, and S. I. Allakhverdiev. 2021. Crop photosynthesis for the twenty-first century. Photosynthesis Research 150(1):1–3.
- Campbell, B. J., A. F. Berrada, and C. Hudalla. 2019. Genotype x environment interactions of industrial hemp cultivars highlight diverse responses to environmental factors. Agrosystems, Geosciences & Environment 2(1):180057.
- Caplan, D., M. Dixon, and Y. Zheng. 2019. Increasing inflorescence dry weight and cannabinoid content in medical cannabis using controlled drought stress. HortScience 54(5):964–969.
- Chandra, S., H. Lata, I. A. Khan, and M. A. Elsohly. 2008. Photosynthetic response of Cannabis sativa L. to variations in photosynthetic photon flux densities, temperature and CO2 conditions. Physiology and Molecular Biology of Plants 14(4):299–306.
- Chandra, S., H. Lata, I. A. Khan, and M. A. ElSohly. 2011. Temperature response of photosynthesis in different drug and fiber varieties of Cannabis sativa L. Physiology and Molecular Biology of Plants 17(3):297–303.
- Chandra, S., H. Lata, Z. Mehmedic, I. A. Khan, and M. A. ElSohly. 2015. Light dependence of photosynthesis and water vapor exchange characteristics in different high Δ9-THC yielding varieties of Cannabis sativa L. Journal of Applied Research on Medicinal and Aromatic Plants 2(2):39–47.
- Chen, X., H. Deng, J. A. Heise, D. P. Puthoff, N. Bou-Abboud, H. Yu, and J. Peng. 2021. Contents of cannabinoids in hemp varieties grown in Maryland. ACS Omega 6(47):32186–32197.
- Cherney, J. H., and E. Small. 2016. Industrial hemp in North America: production, politics and potential. Agronomy 6(4):58.
- Chiluwal, A., S. S. Sandhu, H. Sandhu, M. Irey, F. Johns, and R. Sanchez. 2023. Cannabidiol industrial hemp growth, biomass, and temporal cannabinoids accumulation under different planting dates in southern Florida. Agrosystems, Geosciences & Environment 6(1):e20347.
- Cockson, P., H. Landis, T. Smith, K. Hicks, and B. E. Whipker. 2019. Characterization of nutrient disorders of Cannabis sativa. Applied Sciences 9(20):4432.
- Cockson, P., M. Schroeder-Moreno, P. Veazie, G. Barajas, D. Logan, M. Davis, and B. E. Whipker. 2020. Impact of phosphorus on Cannabis sativa reproduction, cannabinoids, and terpenes. Applied Sciences 10(21):7875.
- Duursma, R. A. 2015. Plantecophys-an R package for analyzing and modelling leaf gas exchange data. PLOS ONE 10(11):e0143346.
- Eichhorn Bilodeau, S., B. S. Wu, A. S. Rufyikiri, S. MacPherson, and M. Lefsrud. 2019. An update on plant photobiology and implications for cannabis production. Frontiers in Plant Science 10:296.
- Evans, J. R. 1989. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 78(1):9–19.
- Farquhar, G. D., S. V. von Caemmerer, and J. A. Berry. 1980. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149(1):78–90.
- Farquhar, G. D., and T. D. Sharkey. 1982. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology 33(1):317–345.
- Gilbert, M. E., M. A. Zwieniecki, and N. M. Holbrook. 2011. Independent variation in photosynthetic capacity and stomatal conductance leads to differences in intrinsic water use efficiency in 11 soybean genotypes before and during mild drought. Journal of Experimental Botany 62(8):2875–2887.
- Gregory, L.M., A. M. McClain, D. M. Kramer, J. D. Pardo, K. E. Smith, O. L. Tessmer, B. J. Walker, L. G. Ziccardi, and T. D. Sharkey. 2021. The triose phosphate utilization limitation of photosynthetic rate: out of global models but important for leaf models. Plant, Cell & Environment 44(10):3223–3226.
- Grossiord, C., T. N. Buckley, L. A. Cernusak, K. A. Novick, B. Poulter, R. T. Siegwolf, J. S. Sperry, and N. G. McDowell. 2020. Plant responses to rising vapor pressure deficit. New Phytologist 226(6):1550–1566.
- Hatfield, J. L., and C. Dold. 2019. Water-use efficiency: advances and challenges in a changing climate. Frontiers in Plant Science 10:103.
- Jenkins, M. W. 2021. Cannabis sativa L. response to narrow bandwidth UV and the combination of blue and red light during the final stages of flowering on leaf-level gas-exchange parameters, secondary metabolite production, and yield. Agricultural Sciences 12:1414–1432.
- Jenkins, M. W., and C. B. Livesay. 2021. Photosynthetic performance and potency of Cannabis sativa L. grown under LED and HPS illumination. Agricultural Sciences 12:293–304.
- Kumarathunge, D. P., B. E. Medlyn, J. E. Drake, A. Rogers, and M. G. Tjoelker. 2019. No evidence for triose phosphate limitation of light‐saturated leaf photosynthesis under current atmospheric CO2 concentration. Plant, Cell & Environment 42(12):3241–3252.
- Leakey, A. D., J. N. Ferguson, C. P. Pignon, A. Wu, Z. Jin, G. L. Hammer, and D. B. Lobell. 2019. Water use efficiency as a constraint and target for improving the resilience and productivity of C3 and C4 crops. Annual Review of Plant Biology 70:781–808.
- LiCor Biosciences. 2022. Complementary leaf physiology measurements: the LI-600 and LI-6800. Application Note #979-19769.
- Linder, E. R., S. Young, X. Li, S. Henriquez Inoa, and D. H. Suchoff. 2022. The effect of harvest date on temporal cannabinoid and biomass production in the floral hemp (Cannabis sativa L.) cultivars BaOx and Cherry Wine. Horticulturae 8(10):959.
- Lombardozzi, D. L., N. G. Smith, S. J. Cheng, J. S. Dukes, T. D. Sharkey, A. Rogers, R. Fisher, and G. B. Bonan. 2018. Triose phosphate limitation in photosynthesis models reduces leaf photosynthesis and global terrestrial carbon storage. Environmental Research Letters 13(7):074025.
- Long, S. P., and C. J. Bernacchi. 2003. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. Journal of Experimental Botany 54(392):2393–2401.
- Long, S. P., X. G. Zhu, S. L. Naidu, and D. R. Ort. 2006. Can improvement in photosynthesis increase crop yields? Plant, Cell & Environment 29(3):315–330.
- Lydon, J., A. H. Teramura, and C. B. Coffman. 1987. UV‐B radiation effects on photosynthesis, growth and cannabinoid production of two Cannabis sativa chemotypes. Photochemistry and Photobiology 46(2):201–206.
- Malceva, M., M. Vikmane, and V. Stramkale. 2011. Changes of photosynthesis-related parameters and productivity of Cannabis sativa under different nitrogen supply. Environmental and Experimental Biology 9:61–69.
- Malone, T., and K. Gomez. 2019. Hemp in the United States: a case study of regulatory path dependence. Applied Economic Perspectives and Policy 41(2):199–214.
- Mark, T., J. Shepherd, D. Olson, W. Snell, S. Proper, and S. Thornsbury. 2020. Economic viability of industrial hemp in the United States: a review of state pilot programs. USDA. Economic Research Service, EIB-217.
- Meter Group, Inc. USA. 2018. Soil Electrical Conductivity – A Guide to Measurements. Application Note #18190-02.
- Miner, G. L., and W. L. Bauerle. 2019. Seasonal responses of photosynthetic parameters in maize and sunflower and their relationship with leaf functional traits. Plant, Cell & Environment 42(5):1561–1574.
- Moher, M., D. Llewellyn, M. Jones, and Y. Zheng. 2022. Light intensity can be used to modify the growth and morphological characteristics of cannabis during the vegetative stage of indoor production. Industrial Crops and Products 183:114909.
- Moore, C. E., K. Meacham-Hensold, P. Lemonnier, R. A. Slattery, C. Benjamin, C. J. Bernacchi, T. Lawson, and A. P. Cavanagh. 2021. The effect of increasing temperature on crop photosynthesis: from enzymes to ecosystems. Journal of Experimental Botany 72(8):2822–2844.
- Murovec, J., J. J. Eržen, M. Flajšman, and D. Vodnik. 2022. Analysis of morphological traits, cannabinoid profiles, THCAS gene sequences, and photosynthesis in wide and narrow leaflet high-cannabidiol breeding populations of medical cannabis. Frontiers in Plant Science 13:786161.
- Pennsylvania Department of Agriculture. 2022 Hemp Varieties of Concern & Prohibited Varieties. Available from: https://www.agriculture.pa.gov/Plants_Land_Water/hemp/Documents
- Peterson, A. G., J. T. Ball, Y. Luo, C. B. Field, P. B. Reich, P. S. Curtis, K. L. Griffin, C. A. Gunderson, R. J. Norby, D. T. Tissue, M. Forstreuter, A. Rey, and C. S. Vogel. 1999. The photosynthesis–leaf nitrogen relationship at ambient and elevated atmospheric carbon dioxide: a meta‐analysis. Global Change Biology 5(3):331–346.
- Reich, P. B., D. S. Ellsworth, M. B. Walters, J. M. Vose, C. Gresham, J. C. Volin, and W. D. Bowman. 1999. Generality of leaf trait relationships: a test across six biomes. Ecology 80(6):1955–1969.
- Rodriguez-Morrison, V., D. Llewellyn, and Y. Zheng. 2021. Cannabis yield, potency, and leaf photosynthesis respond differently to increasing light levels in an indoor environment. Frontiers in Plant Science 12:456.
- Rogers, A., D. P. Kumarathunge, D. L. Lombardozzi, B. E. Medlyn, S. P. Serbin, and A. P. Walker. 2021. Triose phosphate utilization limitation: an unnecessary complexity in terrestrial biosphere model representation of photosynthesis. New Phytologist 230:17–22.
- Saathoff, A. J., and J. Welles. 2021. Gas exchange measurements in the unsteady state. Plant, Cell & Environment 44(11):3509–3523.
- Saloner, A., and N. Bernstein. 2020. Response of medical cannabis (Cannabis sativa L.) to nitrogen supply under long photoperiod. Frontiers in Plant Science 11:572293.
- Shiponi, S., and N. Bernstein. 2021. The highs and lows of P supply in medical cannabis: effects on cannabinoids, the ionome, and morpho-physiology. Frontiers in Plant Science 12:657323.
- Sperry, J. S. 2000. Hydraulic constraints on plant gas exchange. Agricultural and Forest Meteorology 104(1):13–23.
- Stinziano, J. R., P. B. Morgan, D. J. Lynch, A. J. Saathoff, D. K. McDermitt, and D. T. Hanson. 2017. The rapid A–Ci response: photosynthesis in the phenomic era. Plant Cell and Environment 40(8):1256–1262. https://doi.org/10.1111/pce.12911
- Stromquist, A., and H. Sundelius. 1969. Stratigraphy of the Albemarle Group of the Carolina Slate Belt in central North Carolina. Geological Survey Bulletin 1274-B. U.S Department of the Interior, Washington, D.C., USA.
- Suchoff, D., J. Davis, M. Bloomquist, and M. Short. 2022. Understanding the relationship between THC and CBD in hemp. AG-926. North Carolina State Cooperative Extension Service, Raleigh, NC, USA.
- Sunoj Valiaparambil Sebastian, J., X. Dong, C. Trostle, H. Pham, M. V. Joshi, R. W. Jessup, M. D. Burrow, an dt. L. Povin. 2023. Hemp agronomy: current advances, questions, challenges, and opportunities. Agronomy 13(2):475.
- Tang, K., P. C. Struik, S. Amaducci, T. J. Stomph, and X. Yin. 2017. Hemp (Cannabis sativa L.) leaf photosynthesis in relation to nitrogen content and temperature: implications for hemp as a bio‐economically sustainable crop. GCB Bioenergy 9(10):1573–1587.
- Tang, K., A. Fracasso, P. C. Struik, X. Yin, and S. Amaducci. 2018. Water-and nitrogen-use efficiencies of hemp (Cannabis sativa L.) based on whole-canopy measurements and modeling. Frontiers in Plant Science 9:951.
- Taylor, S. H., and S. P. Long. 2019. Phenotyping photosynthesis on the limit–a critical examination of RACiR. The New Phytologist 221(2):621–624.
- Trancoso, I., G. A. de Souza, P. R. dos Santos, K. D. dos Santos, R. M. D. S. N. de Miranda, A. L. P. M. da Silva, D. Z. Santos, I. F. García-Tejero, and E. Campostrini. 2022. Cannabis sativa L.: crop management and abiotic factors that affect phytocannabinoid production. Agronomy 12(7):1492.
- Veazie, P., P. Cockson, D. Kidd, and B. Whip. 2021. Elevated phosphorus fertility impact on Cannabis sativa ‘BaOx’growth and nutrient accumulation. International Journal of Innovative Science, Engineering & Technology 8:345–351.
- Walker, A. P., A. P. Beckerman, L. Gu, J. Kattge, L. A. Cernusak, T. F. Domingues, J. C. Scales, G. Wohlfahrt, S. D. Wullschleger, and F. I. Woodward. 2014. The relationship of leaf photosynthetic traits—Vcmax and Jmax—to leaf nitrogen, leaf phosphorus, and specific leaf area: a meta‐analysis and modeling study. Ecology and Evolution 4(16):3218–3235.
- Wong, S. C., I. R. Cowan, and G. D. Farquhar. 1979. Stomatal conductance correlates with photosynthetic capacity. Nature 282(5737):424–426.
- Wullschleger, S. D. 1993. Biochemical limitations to carbon assimilation in C3 plants – a retrospective analysis of the A/Ci curves from 109 species. Journal of Experimental Botany 44(5):907–920.