

REVIEW PAPER
Daniel Airola*
Conservation Research and Planning, Sacramento, CA, USA![]() https://orcid.org/0000-0001-6725-4230
https://orcid.org/0000-0001-6725-4230
*Corresponding Author: d.airola@scglobal.net
Published 28 March 2024 • doi.org/10.51492/cfwj.110.2
Abstract
The purple martin (Progne subis) has declined substantially in numbers and geographic range since the 1980s. Surveys and monitoring are needed to better understand the status of the species, in order to set conservation priorities and direct actions. Purple martins in California often breed in habitats that differ from those in other areas of the species’ range. The species’ sparse distribution, colonial nature, and frequent use of inaccessible nests sites require specialized methodologies to determine suitable nesting habitat, occupancy, numbers, and reproductive success. I present a range of methods available to monitor nesting purple martins in the variety of nesting substrates in which they nest in California. These methods should improve efficiency and accuracy of surveys and provide comparable results that can be used to evaluate regional or statewide trends.
Key words: occupancy, population size, productivity, Progne subis, purple martin, survey methods
| Citation: Airola, D. 2024. Survey methods for the purple martin in California. California Fish and Wildlife Journal 110:e2. |
| Editor: Matthew Toenies, Office of Cannabis |
| Submitted: 2 November 2023; Accepted: 11 December 2023 |
| Copyright: ©2024, Airola. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Competing Interests: The author has not declared any competing interests. |
Introduction
The purple martin (Progne subis)is designated as a Species of Special Concern by the California Department of Fish and Wildlife because of substantial declines in its numbers and geographic range (Shuford and Gardali 2008; Airola and Williams 2008). Purple martins in California are of the western subspecies (P. s. arboricola), which is scattered across western North America west of the Rocky Mountains and north of the Sonoran Desert (Brown et al. 2021). The western subspecies is much less abundant than the well-known eastern purple martin (P. s. subis), which depends almost entirely on human-supplied nest boxes (Tautin et al. 2009), and probably less abundant than the southwestern subspecies (P. s. hesperia) that nests mainly in saguaro cacti (Carnegiea gigantea) in the Sonoran Desert (Brown et al. 2021).
The purple martin population decline in California occurred mainly since the 1980s due to nest site competition by the non-native European starling (Sturnus vulgaris) following its arrival as a breeding species in California (Airola and Grantham 2003; Airola and Williams 2008). Continuing recent declines have been attributed to effects of insecticides on insect prey and martin reproduction and on construction disturbance in and around occupied bridges, overpasses, and elevated freeways (Airola et al. 2014; Airola and Kopp 2018; Airola 2020).
Purple martins are secondary cavity-nesters (i.e., non-excavators) that breed colonially in a variety of nesting substrates in California (Airola and Williams 2008). Most nests are in holes in snags (standing dead trees) and live trees, but they nest extensively in bridges and utility structures, and very locally in nest boxes (Airola et al. 2018). The species formerly nested extensively in buildings but has not done so (with very few exceptions) since the starling’s arrival (Airola and Grantham 2003). Use of snags and live trees was formerly more widespread but is now limited to coastal regions, montane conifer forests, and the Tehachapi Mountains where starlings are less abundant.
Martins are highly vocal and forage and nest in open areas, so they are relatively easy to detect. Most nesting substrates (except nest boxes), however, pose substantial access limitations for conducting nest checks to determine reproduction. Thus, specialized methods have been developed to allow determination of the various breeding parameters that may be possible or practical to obtain to meet survey goals in a variety of local settings, including determining occurrence, nesting use, and nesting success (Airola and Grantham 2003; Airola 2009, 2020; Airola et al. 2018).
Purple martin surveys are needed to identify occupied breeding sites for conservation purposes and to monitor population status and breeding success. Different survey methods may be applied depending on survey goals. This document provides comprehensive guidance on how to conduct the full range of surveys in various nest substrates to meet various survey goals.
Species-specific Considerations in Designing and Conducting Surveys
Many species-specific factors must be considered in conducting reliable surveys for purple martins. Martins are scattered widely at low densities across much of their range; large areas that appear suitable for the species are not occupied. The species is colonial, so social attraction has a substantial influence over occurrence. The concentration of birds within colonies, however, can make them easier to miss in larger-scale surveys. The timing of arrival at breeding sites from migration and the timing of breeding activities varies substantially with temperature, and thus latitude, elevation, and the extent of coastal influence. For example, arrival and breeding activities in warmer Sacramento occur a month earlier than along the cooler Mendocino Coast (Airola 2009; see Colony Dectection below).
Male Purple Martins are reported to typically return to colonies earlier than females (Brown et al. 2021), but no difference in arrival dates has been found in Sacramento in five colonies monitored over seven years (D. Airola, unpub. data.). Martins also have a complex age structure with subadult (“second-year” or 1-year-old) males showing a distinctive plumage that is more similar to females than to adult (“after second-year”) males. The timing of return to breeding sites from migration is age-specific, with subadults (1-year-old birds), some of which breed in their first year (Airola 2009), returning up to six weeks later than adults. Breeding purple martins spend much of their time within their nest cavities or foraging away from the colony site, which reduces their detectability. Snags used for nesting are subject to decay and loss, and thus despite strong philopatry, martins are periodically required to relocate to new sites, thereby inhibiting long-term monitoring efforts. Conversely, several martin characteristics facilitate their survey. They are loud and fly in the open, so are easy to detect. Their strong philopatry allows them to be readily re-found in bridges and other permanent structures for multi-year monitoring.
Colony Detection
An initial goal of more extensive surveys is to determine if purple martins are present at a site or are likely to occur.
Determining Potential for Occurrence
Determining if a site has potential for occurrence of purple martins can be challenging because many types of sites can be suitable, and areas that appear to have suitable habitat conditions are often unoccupied. Therefore, past records can be valuable in determining a site’s potential for present occurrence.
Records of historical occurrence (i.e., before the mid-1990s) were summarized comprehensively by Williams (1998). Many of these records, however, precede the arrival and population increase of the European Starling in the 1970s, and thus many of these identified sites are no longer occupied. More recent information sources include eBird, various Breeding Bird Atlases (Appendix (PDF)), and studies conducted in various areas of the state (Table 1).
Table 1. Locations with recent (>1990) published studies of purple martins in California.
| Location | Nesting Substrate | Reference |
| Lake Britton, Shasta Co. | Lakeshore snags | Airola 2009 |
| Shasta Lake, Shasta Co. | Flooded snags | Lindstrand 2008 |
| Shelter Cove, Humboldt Co. | Snags, nest boxes, utility poles, buildings | Kostka et al. 2008; Elwood et al. 2009; Airola et al. 2018 |
| Mendocino Coast, Mendocino Co. | Bridges, snags | Airola 2009 |
| Red Hills Road, Lake Co. | Electrical transmission poles | Woodward and Woodward 2005; Airola 2009 |
| Sacramento, Sacramento Co. | Bridges | Airola and Grantham 2003, Airola 2020; Airola and Kopp 2021 |
| Dry Creek and Yokohl Valley, Tulare Co. | Electrical transmission towers | Sylvester and Airola 2010 |
| Big Sur Town, Monterey Co | Snags, bridges | Airola 2009 |
| Atascadero and Santa Margarita area, San Luis Obispo Co. | Trees, snags | Airola 2009 |
| Tejon Ranch, Kern Co. | Trees | Williams 2002; White et al. 2011 |
eBird is the most up-to-date information source on current occurrences, as purple martins are rare enough that they are readily sought out and recorded by birders through much of their range in California. eBird data can provide valuable information on occurrence, relative numbers, and evidence of breeding, especially in areas that are visited frequently. Many eBird records, however, are scattered, recorded opportunistically, and lack details on evidence of breeding (versus migratory or transitory occurrence). Also, large areas of private lands are inaccessible to most birders, so eBird records are mostly restricted to public lands and along public roads. Therefore, while eBird can be useful in documenting many breeding occurrences, a lack of previous records is not a sufficient basis on which to conclude a site is unoccupied.
Verifying Site Suitability
Purple martins are widely but locally distributed in forest and woodland areas at low to intermediate elevations in California (Airola and Williams 2008). Recent review of eBird data shows that most martins nest in forests throughout the Coast Ranges, and breeding in the Sierra Nevada now appears to be limited to a few areas in the north (D. Airola, unpub. data). Scattered populations also occur in interior low-to-mid elevation areas. Much apparently suitable habitat, however, is not occupied, suggesting a need for more detailed modelling of determinants of occurrence such as done elsewhere by Williams et al. (2020).
Areas targeted for survey can be evaluated for suitability before the survey period. Suitability is mostly based on the presence of suitable nesting habitat. Sites that may be suitable for martin occurrence include snags, live trees, utility poles, bridges (including elevated freeways and overpasses >70 m long), and unusually, in buildings, transmission towers, elevated pipes, and underground lava tubes. All suitable nest sites are in open areas that provide flight access. Martin accessibility to nesting substrates should be evaluated in the field, as access to sites used in the past may become obstructed by vegetation growth (Airola 2009) or building construction (Airola and Grantham 2003). Nest sites are often located on slopes or ridge tops (Airola and Williams 2008; White et al. 2011), usually with a water body nearby (for drinking and insect production). Areas in which no nesting habitat is present or accessible to birds need not be surveyed.
Selecting the Survey Period
The timing of the purple martin nesting season varies substantially across California with latitude and elevation likely due to sensitivity to temperature and resulting insect abundance. While the general outlines of the timing are known across the state, and are known in detail in some areas, applied judgement is needed to select the appropriate time for surveys in much of the state. As shown in Table 2, the general pattern is that nesting occurs later in areas further north, at higher elevation, and closer to the coast.
Table 2. Known average timing of nesting activities at different latitudes and elevations in California. Sites listed from south to north.a
| Location | Region | County | Elevation (m) | Annual First Arrival | Nest Building | Incubating | Nestlingb | Pre-fledgling Youngc |
| Nojoqui Falls Park | Tehachapi Mts/South Coast | Santa Barbara | 300 | — | 19 May | — | 29 Jun | — |
| Tejon Ranch | Tehachapi Mts/South Coast | Kern | 300–500 | — | — | — | 30 Jun | — |
| Sacramento | Central Valley and Interior Coast Range | Sacramento | 5 | 18 Mar | 25 May | 10 Jun | 24 Jun | 1 Jul |
| Pope Valley | Central Valley and Interior Coast Range | Napa | 225 | 18 Mar | — | — | — | — |
| Red Hills Rd | Central Valley and Interior Coast Range | Lake | 650 | 20 Mar | 15 May | — | 30 Jun | 5 Jul |
| Mendocino Coast | Northwest Coast | Mendocino | 10 | 14 Apr | 17 Jun | — | — | — |
| Shelter Cove | Northwest Coast | Humboldt | 35 | 5 Apr | 27 Jun | 29 Jun | 10 Jul | 22 Jul |
| Shasta Lake | Northwest Interior | Shasta | 100 | 3 Apr | 7 Jun | — | 25 Jun | 1 Jul |
| Lake Britton | Northwest Interior | Shasta | 900 | — | 15 Jun | — | 30 Jun | 5 Jul |
| Lava Beds National Monument | Northwest Interior | Siskiyou | 1,600 | — | — | — | — | 18 Jul |
a Sources: Purple Martin Conservation Association (https://www.purplemartin.org/research/8/scout-arrival-study/), Airola 2009, 2020, unpub data; White et al. 2011; ebird.org.
b Nestling carrying food or fecal sac
c Heard or seen in nest
Determining Occupancy
I define occupancy as the presence of birds that are breeding or attempting to breed at a site. Although martins are relatively easy to detect through vocalization and direct observation, certain conditions increase activity and thus the likelihood of detection at occupied sites. They are generally not active at temperatures below about 12o C (53o F) or above 35o C (95o F). When conditions are cool, generally below 14o C (58 o F), martins either remain inside nests or fly to water bodies where warmer temperatures maintain higher insect activity and are thus less detectable. Less is known about behavior during very high temperatures, which suppresses insect activity, but presumably birds remain within nest sites. Martins are most active around colony sites during the several hours after sunrise, when courtship, active feeding, and food deliveries to young occur most frequently.
The nesting stage strongly affects martin activity and thus detection (Table 3). Martins are highly active during the settling/courtship and nest building stages, and then activity decreases substantially during incubation. After young hatch, activity continues increasing as food demand by nestlings increases. Following fledging, martin families typically roost at night within nest cavities (Airola and Grantham 2003; Airola 2020; Brown et al. 2021), and adults move young away from the nesting colony to feed them during the daytime. Thus, activity at colony sites is low during this period, except when leaving the colony in the morning and returning in the evening. Martins then abandon nesting areas generally within several weeks of fledging young.
The later arrival and breeding by subadult birds also complicates selection of survey times. Determining occupancy at sites occupied only by subadults (which are often newly occupied) requires extending surveys later in the nesting period, to match periods of higher activity and detection.
Incorporating all the factors affecting detectability, surveys for occupancy (and to estimate populations, see Determining Numbers below) should be tailored to periods of high activity. I developed and tested a method that was successful that requires a minimum of three surveys of potential nest sites: one during courtship/nest site selection and nest building, and two during the nestling period (Airola 2009). Of course, more surveys provide better information on occupancy.
Table 3. Levels of purple martin activity and thus and detectability during various nesting stages
| Nesting Stage | Duration (days)a | Activity Level |
|---|---|---|
| Courtship and nest site selection | 30–65 | High |
| Nest-building | 2–10 | High |
| Egg-laying | 4–6 | Moderate |
| Incubation | 15–19 | Low |
| Nestling | 26–33 | High |
| Fledgling | 5–14b | Lowc |
| Total | 82–147 | — |
a Sources: Airola 2020; Brown et al. 2021; D. Kopp pers. comm.; D. Airola unpub. data
b Duration for this stage is the period when fledglings typically return to nest sites to roost overnight.
c Activity is seldom detected during the daytime during this period because adults typically lead fledglings away from the nest site in the early morning and return with them to roost at dusk. Thus, activity and detection are higher during morning departure and evening arrival.
Determining Numbers of Breeding Adults
Purple martin nests are generally inaccessible for direct examination because of their placement within trees, bridges, and utility poles. Next boxes can be readily examined, but in recent history, boxes have only been used at two sites in California (Airola et al. 2018, D. Airola, unpub. data). Because of the difficulties of surveying for purple martins, estimating numbers requires use of inductive methods. Several methods exist that have different levels of reliability and precision.
Direct Counts
As noted under Determining Occupancy, the total number of adults at a purple martin breeding colony typically cannot be directly observed because at any time some birds are in holes and others may be away from the colony. Also, use of the more intensive hole mapping method (see below) may be infeasible where topography, visibility, and access are challenging. Therefore, it is highly likely that most of the numbers of birds reported anecdotally from single visits, such as in eBird, are underestimates of true numbers.
To address these challenges, I previously evaluated the use of direct counts as an index to the total number of birds breeding at a colony sites (Airola 2009). For this comparison, I used Sacramento bridge colonies with known numbers of martins, determined from hole use and behavior. This analysis showed that the average of three direct counts conducted over the nesting season generally detected 50% of the total breeding population at colonies. Therefore, conducting counts on three dates spread over the nesting season and multiplying the average by two provides a reasonable, if rough, estimate of the colony size at bridges. Comparison of numbers at occupied colonies determined through hole use mapping to numbers counted across Northern California (Airola 2009) yielded different multipliers for counts at bridges (2.17, n = 14), snags and utility poles (1.41, n = 8), and caves (1.12, n = 4), apparently reflecting different martin detectability in these settings. Thus, applying these multipliers to counts of martins nesting in different substrates is recommended.
Playing recorded vocalizations, especially purple martin alarm calls, may be useful in attracting more of the resident birds and making them more visible at colony sites for counting. Playback has not been reported to be used for this purpose, but rather mainly to attract martins to colonize new sites (Doughty and Fergus 2002; Airola et al. 2018). The technique deserves further evaluation but would require recalibrating the 50% detection proportion applied to non-playback counts.
Another approach to estimate a minimum number of breeders is to count the number of adult males, whose plumage make them most easily recognizable. The number of adult males multiplied by two provides a minimum number but does not account for adult males that may not have been visible during counts, or breeding subadult males, which are recognizable typically only by those with considerable field experience (Pyle 1997; Hill 2002; Brown et al. 2021). Subadult males have been found to comprise an average of 8% of breeding males in Sacramento bridges, which could be used as a general guide to adjust the estimates, but the annual proportions of breeding subadults in populations can vary substantially, from 0–22% (Airola 2020; Airola and Kopp 2021, 2023).
Mapping Hole-use and Recording Diagnostic Breeding Behaviors
The most reliable survey method to determine martin colony size combines quantifying the use of different nest sites and recording behaviors that indicate breeding use. This method has been used and evaluated at Sacramento colonies since the 1990s, where it is considered accurate in providing a complete population census (Airola 2020). It also has been employed effectively in surveying colonies in a wide variety of nesting substrates during surveys across Northern California (Airola 2009).
The hole-use mapping method employs repeated visits to nesting colonies at key times during the nesting season to map martin use of individual holes and record behaviors that are diagnostic of breeding (i.e., nest building, carrying food to nests, removing nestling fecal sacs, or sounds or observations of begging young). Mapping hole use and recording behavior over multiple visits is required to separate nest sites from sites that are being “explored” by as nest sites by after-second-year birds (especially early in the season) or by later-arriving subadult birds as part of courtship and pair-bonding.
Maps are created with suitable nest sites individually identified (i.e., with unique alphanumeric codes) so that hole entries and exits can be recorded and behaviors noted (Fig. 1). Over the course of a season, cumulative hole entries and exits and associated breeding behaviors (Fig. 1) are summarized in spreadsheets (Fig. 2). Examination of the pattern of use of a potential nest site and the display of diagnostic breeding behaviors can clearly indicate which sites are confirmed as breeding sites used by pairs. In cases where conditions do not allow observation of breeding behaviors, I and colleagues typically count a pair as actively breeding if we observe it entering a suitable nest hole on at least two dates separated by at least two weeks during the height of the nesting season (Fig. 2).

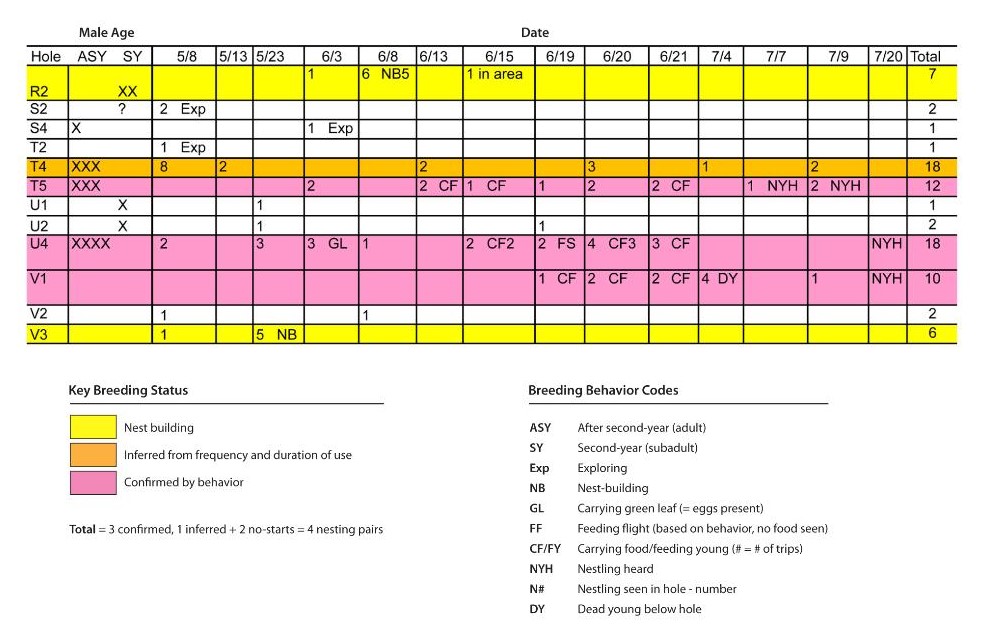
Applying survey methods that require multiple visits to sites can be challenging in remote locations. Airola (2009) developed a survey method to employ counts and hole mapping over three visits: one during the early nesting period and two visits during the late period. The method balances the quality of information with efficiency and provides a standardized method that is comparable among sites. To be effective, survey timing needs to match the breeding periods of martins at different latitudes, elevations, and climates (Table 2).
Determining Nesting Success
Nesting success can be determined with different levels of precision, depending on the type of nest site and access for observation. The two main measures are breeding success, which indicates whether or not a nest was successful in producing any fledglings, and reproductive success (or productivity), which determines the number of fledglings produced by nesting pairs. Understanding the limitations in accurately determining breeding success and reproductive success are important to understand in applying methods.
Breeding success may be the best indicator of nesting success that can be determined at many sites that are inaccessible for direct nest viewing, such as in bridges, snags, or utility poles. Whether nesting was successful can be determined by a combination of observing the presence of young near fledging age within the nest cavity or observing fledglings around the nest site, especially when leaving in the morning or returning at night, which typically happens for 1–2 weeks after fledging occurs (Airola 2020; Brown et al. 2021). The duration of activity at a nest site also can indicate whether a site is successful (i.e., compared to timing of stages; Table 3). For example, nestlings typically can be heard calling in the nest for a week or so before fledging. Also, older young often may perch at the entrance hole to wait for food deliveries from adults. The presence of older young indicates that a nest is likely to be successful.
Determining reproductive success through direct observation is possible only in a few circumstances. Nest-boxes, of course, can generally be directly examined to count young nearing fledgling age, but martins use nest boxes only at a few sites in California (Airola et al. 2018). In some cases, young can be counted returning to holes to roost at dusk, but where multiple pairs occur at a site it can be challenging separating different pairs as they circle around colonies under low light conditions before entering their nest sites to roost. Counts of older nestlings in nest holes awaiting food deliveries can indicate a minimum number of likely fledglings, but other nestlings may remain out of sight in the nest hole.
Some snags and poles may be directly accessible using ladders and periscopes or mirrors to see into holes, but snags used by martins are often rotten and should not be used to support a ladder. Examination of nests using pole mounted cameras inserted into nest holes can be feasible for nests in some utility poles, snags, and bridges. Use of poles in bridges, however, can be difficult because the nesting chambers are large, and they are often filled with construction debris that obscures visibility. Also, by age 20 days, nestlings often react to the presence of the camera by moving away and hiding out of sight (Airola et al. 2008). Because of these issues and low visibility inside bridges, counts within bridges are often questionable (Airola 2020). Use of tall pole-mounted cameras at bridges and utility poles also can be hazardous, due to traffic and electrocution, respectively. Finally, using cameras to monitor purple martin nests may require permits from the California Department of Fish and Wildlife.
Documenting Results
Survey results should be publicly documented to assist in ongoing efforts to understand the purple martin’s status and population trends in California. eBird (ebird.org) offers the easiest way to record observations publicly. The California Natural Diversity Data Base (https://wildlife.ca.gov/Data/CNDDB) contains fewer purple martin records than eBird, due to a more arduous process for entering data, but is a worthwhile repository because it is used extensively in conducting impact assessments. In addition to occurrence, observers should report details of behavior, including observation of any diagnostic breeding behaviors and nesting substrates. More extensive surveys should be prepared for publication in local or regional journals, such as Central Valley Birds, Western Birds, or California Fish and Wildlife Journal.
Literature Cited
- Airola, D. A. 2009. Status of purple martin populations in northern California: results of a pilot study to develop and apply a survey methodology. U. S. Fish and Wildlife Service, Sacramento Field Office, CA, USA.
- Airola, D. A. 2020. Life Under the Fast Lane: Ecology and Conservation of the Bridge-Nesting Purple Martins in Urban Sacramento. Special Publication No. 1, Central Valley Bird Club, Sacramento, CA, USA.
- Airola, D. A., B. Cousens, and D. Kopp. 2014. Accelerating decline of the Sacramento purple martin breeding population in 2014: what are the possible causes? Central Valley Bird Club Bulletin 17:12–22.
- Airola, D. A., and J. Grantham. 2003. Purple martin population status, nesting habitat characteristics, and management in Sacramento. Western Birds 34:235–251.
- Airola, D. A., and D. Kopp. 2018. Another substantial decline in the Sacramento purple martin nesting population in 2018: the role of construction disturbance and future threats. Central Valley Bird Club Bulletin 21:75–88.
- Airola, D. A., and D. Kopp. 2021. Sacramento purple martins in 2021: results of breeding surveys and conservation actions. Central Valley Birds 24:43–49.
- Airola, D. A., and D. Kopp. 2023. Customized protection measures allow purple martins to nest successfully during freeway construction. Central Valley Birds 26:109–122.
- Airola, D. A., D. Kopp, and K. Thomas. 2008. Population status, reproduction, and mortality of purple martins in Sacramento during 2007. Central Valley Bird Club Bulletin 11:25–36.
- Airola, D. A., S. Kostka, and C. Elwood. 2018. Attempts to establish colonies of the purple martin in nest boxes in California: first success and evaluation of failures. Western Birds 49:113–125.
- Airola, D. A., and B. D. C. Williams 2008. Purple martin (Progne subis). Pages 293–299 in W. D. Shuford and T. Gardali, editors. California Bird Species of Special Concern: A Ranked Assessment of Species, Subspecies, and Distinct Populations of Birds of Immediate Conservation Concern in California. Studies of Western Birds 1. Western Field Ornithologists, Camarillo, CA and California Department of Fish and Game, Sacramento, CA, USA. https://nrm.dfg.ca.gov/FileHandler.ashx?DocumentID=10417&inline
- Allen, H., D. Hatch, D. Humple, L. Lewis, and W. Wiskes. Marin County Breeding Bird Atlas II: Annual Report for 2023. Marin Audubon Society, Mill Valley, CA, USA.
- Berner, M. B., Grummer, R. Leong, and M. Rippey. 2003. Breeding Birds of Napa County, California, an Illustrated Atlas of Nesting Birds. Napa-Solano Audubon Society, Vallejo, CA, USA.
- Bousman, W. G. 2007. Breeding Bird Atlas of Santa Clara County, California. Santa Clara Valley Audubon Society, Cupertino, CA, USA.
- Burridge, B. 1995. Sonoma County Breeding Bird Atlas. Madrone Audubon Society, Santa Rosa, CA, USA.
- Brown, C. R., D. A. Airola, and S. Tarof. 2021. Purple martin (Progne subis), version 2.0. In Birds of the World (P. Rodenwald, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.purmar.01
- Doughty, R., and R. Fergus. 2002. The Purple Martin. University of Texas Press, Austin, TX, USA.
- Elwood, C., S. Wickland, S. Kostka, and D. A. Airola. 2009. Success continues for pilot nest box program at Shelter Cove, California. Purple Martin Update 18(2):27–29.
- Hill, J. R. 2002. Tattletails: how to tell the sex and age of purple martins by their undertail coverts. Purple Martin Update 11:16–17.
- Hunter, J. E., D. Fix, G. A. Schmidt, and J. C. Power. 2005. Atlas of the Breeding Birds of Humboldt County, California. Redwood Region Audubon Society, Eureka, CA, USA.
- Kostka, S., D. A. Airola, and G. Switzer. 2008. Recent use of nest boxes by purple martins in Northern California. Purple Martin Update 17(3):13–15.
- Lindstrand, L., III. 2008. Purple martin distribution and nesting habitat at Shasta Lake, California. Western Birds 39:166–170.
- Pandolfino, E. R., L.A. Douglas, T. D. Manolis, and C. Conard. 2021. Sacramento County Breeding Birds: A Tale of Two Atlases and Three Decades of Change. Special Publication 2, Central Valley Bird Club, Davis, CA, USA.
- Pyle, P. 1997. Identification Guide to North American Birds. Part 1. Slate Creek Press, Bolinas, CA, USA.
- Sequoia Audubon Society. 2001. San Mateo County Breeding Bird Atlas. Sequoia Audubon Society, Millbrae, CA, USA.
- Roberson, D., C. Tenney, and S. F. Bailey. 1993. Atlas of Breeding Birds of Monterey County. Monterey Peninsula Audubon Society, Monterey CA, USA.
- Santa Cruz Bird Club. 2021. Santa Cruz County Breeding Bird Atlas II. Annual Report Year 5. Santa Cruz Bird Club, Santa Cruz, CA, USA. https://santacruzbirdclub.org/wp-content/uploads/2021/11/Atlas_Year5_Report_Final.pdf
- Shuford, W. D., and T. Gardali, editors. 2008. California Bird Species of Special Concern: A Ranked Assessment of Species, Subspecies, and Distinct Populations of Birds of Immediate Conservation Concern in California. Studies of Western Birds 1. Western Field Ornithologists, Camarillo, CA and California Department of Fish and Game, Sacramento, CA, USA.
- Shuford, W. D. 1993. The Marin County Breeding Bird Atlas: A Distributional and Natural History of Coastal California Birds. Point Reyes Bird Observatory, Stinson Beach, CA, USA.
- Sylvester, V., and D. A. Airola. 2010. Purple martins nesting in low elevation transmission towers in the San Joaquin Valley, California. Central Valley Bird Club Bulletin 13:69–75.
- Tautin, J., B. Cousens, K. Kostka, S. Kostka, and D. Airola. 2009. Addressing regional declines in purple martin populations. Pages 82–87 in T. Rich, C. Arizmendi, D. Demarest, and C. Thompson, editors. Proceedings of the 4th International Partners in Flight Conference: Tundra to Tropics. Partners in Flight, McAllen, TX, USA.
- Unitt, P. 2004. San Diego Bird Atlas. San Diego Natural History Museum. San Diego, CA, USA.
- White, M. D., E. R. Pandolfino, and A. Jones. 2011. Purple martin survey results at Tejon Ranch in the Tehachapi Mountains of California. Western Birds 42:164–173.
- Williams, B. D. C. 1998. Distribution, habitat associations, and conservation of purple martins in California. Thesis, California State University, Sacramento, CA, USA.
- Williams, B. D. C. 2002. Purple martin in oak woodlands. Pages 323–334 in R. B. Standiford, D. McCreary, and K. L. Purcell, technical coordinators. Proceedings of the Fifth Symposium on Oak Woodlands: Oaks in California’s Changing Landscape. USDA Forest Service General Technical Report PSW-GTR-184, San Diego, CA, USA.
- Williams, J. M., J. Siegrist, and A. M. Wilson. 2020. Support for a relationship between demography and modeled habitat suitability is scale dependent for the purple martin Progne subis. Journal of Animal Ecology. 90:236–366.
- Woodward, D., and B. D. Woodward. 2005. Utility poles provide unusual nest-sites for purple martins in Lake County, California. Purple Martin Update 14:26–29.