

FULL RESEARCH ARTICLE
Vernon C. Bleich*
University of Nevada, Reno, Department of Natural Resources and Environmental Science, 1664 North Virginia Street, Mail Stop 186, Reno, NV 89557, USA![]() https://orcid.org/0000-0002-5016-1051
https://orcid.org/0000-0002-5016-1051
*Corresponding Author: vcbleich@gmail.com
Published 6 June 2024 • doi.org/10.51492/cfwj.110.8
Abstract
Coloration is among the characteristics noted first by observers, and color has a strong influence on how animals are perceived. Abnormal coloration has been reported in a variety of taxa, but less frequently among mammals than other classes of vertebrates. Chromatic disorders occurring among Mammalia include albinism, leucism, piebaldism, melanism, xanthism, erythrism, and isabellinism, but only leucism, melanism, and piebaldism have been confirmed in bighorn sheep (Ovis canadensis); ‘albinism’ has been reported in that taxon, but in the absence confirmatory evidence. Bighorn sheep possessing white, or extremely light-colored pelage, have been described in the literature as albinistic, leucistic, or simply as ‘white’. Isabellinism is a genetic anomaly that manifests as sandy- or cream-colored pelage that appears washed-out and otherwise would be dark in color. Some reports of albinistic, leucistic, or ‘white’ bighorn sheep, however, likely represent animals that were of isabelline coloration. This is the first paper to ascribe this unusual color morph to bighorn sheep, and I (1) provide photographs, references, and other evidence of what appears to be isabellinism in that species; (2) explain why those observing ‘white’ bighorn sheep likely have failed to recognize isabelline coloration; (3) discuss the evolutionary consequences of that anomalous condition in terms of fitness and future adaptation; (4) emphasize the role of what may appear to be meaningless observations in terms of their potential ramifications for understanding or explaining phenomena that heretofore have not been fully explored; and (5) encourage readers to recognize that natural history observations have potentially important implications for the interpretation of adaptation and natural selection, and can provide new insight into prior conclusions or future applications.
Key words: adaptation, albinism, bighorn sheep, chromatic disorder, coloration, fitness, isabellinism, leucism, natural selection, Ovis canadensis, pelage, piebaldism
| Citation: Bleich, V. C. 2024. Isabelline coloration: a heretofore unrecognized chromatic aberration in bighorn sheep. California Fish and Wildlife Journal 110:e8. |
| Editor: Anne Hilborn, Wildlife Branch |
| Submitted: 3 January 2024; Accepted: 22 April 2024 |
| Copyright: ©2024, Bleich. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Competing Interests: The author has not declared any competing interests. |
Introduction
Physical appearance influences the ways in which humans interact with wildlife. For example, coloration has a strong influence on the way wildlife is perceived (Williams et al. 2023), and human responses to unusual or striking coloration range from the spiritual (Morsette 1996; Ferris 2013) to the pragmatic (Fenson 2017). Aside from its significance to evolutionary processes or as anecdotal reports published in descriptive papers, however, color variation has remained little discussed in the mammal literature when compared to other classes of vertebrates, and much remains to be learned (Cotts et al. 2023).
Two factors likely determine color variation in the class Mammalia: pigment regulation that alters the density, distribution, or type of pigments along individual hairs, and pigment patterning, in which the spatial distribution of pigmentation varies across the body; both likely have a distinct basis in genetics but manifest in different parts of developmental pathways (Caro and Mallarino 2022). Melanocytes are the pigment-producing cells in the epidermis, and yield eumelanin and phaeomelanin, which are responsible for black, gray, and dark brown colors, and for red and light brown colors, respectively (Hoekstra 2006). Regulation and production of these pigments involve multiple genes (Lamoreux et al. 2010); see also Flores-Peredo et al. (2021), Caro and Mallarino (2022), Kreling (2023), and Williams et al. (2023).
Abnormal coloration in many species of wildlife has been reported many times and for many decades, but observers frequently have misidentified, miscategorized, or misinterpreted the condition, the causation, or the color in their descriptions (Jones 1920). Numerous investigators have noted that reports of abnormal coloration in mammals or birds have lacked detail, clarity, or consistency of interpretation with respect to chromatic aberrations, and in part reflect (1) a lack of familiarity with terminology associated with irregularities in skin or pelage coloration; (2) the differing traits associated with various pelage anomalies or genetic aberrations; (3) a failure to realize that the causes of such anomalies are multiple and complex; or (4) are often attributed to different animals from a superficial observation (Gratten et al. 2008; Sulem et al. 2008; Coetzer and Grobler 2019; van Deventer et al. 2021; Kalds et al. 2022; Bajpai et al. 2023; Cotts et al. 2023; Kreling 2023).
These shortfalls apply to bighorn sheep (Ovis canadensis), among which uniquely colored individuals have been described variously as albinistic, leucistic, partially albinistic, piebald, melanistic or, simply as “white” (Bleich 2017a, 2017b; and references therein). Such individuals continue to receive special attention, however, as exemplified by Sarian (2019) while describing a female desert bighorn sheep (O. c. nelsoni) as, “… one of the famous white ewes in the Clark Mountains [San Bernardino Co., California] that I have heard about. Everyone was in awe. The white ewe was as white as a Dall (sic) sheep.”
The majority of observations or published references to unusual coloration in bighorn sheep pertain to individuals that reportedly were white in color (Bleich 2017a), but differences exist in expression (Appendix I (PDF)) and causation. Albinistic individuals are characterized by a complete lack of pigmentation in the skin and hair and have pink eyes; the pelage of leucistic individuals (Figs. 1–3) is completely deficient in pigmentation and is white, but they have pigmented skin and eyes that are of normal coloration or are blue (Abreu et al. 2013 for review). Piebaldism occurs when developmental abnormalities result in the absence of mature melanocytes from discrete areas of the body; that condition manifests as white spotting (Spritz 1994) and occurs in bighorn sheep (Bleich 2017a; Blandford 2023c). There is a paucity of reports of melanism (i.e., individuals that exhibit dark, or even black, pelage or appendages; van Grouw 2006) in the mammal literature (Jacques et al. 2008), but it has been described among a variety of mammalian taxa (Baccus and Posey 1999; Hedges et al. 2015; UDWR 2020; Hinton et al. 2022; Bajpai et al. 2023; Cotts et al. 2023) and has been reported at least four times in bighorn sheep (Wilson 1970; O’Connor 1974; Bunch and Nguyen 1982; Conklin Foundation 2022). As with albinism (Fertl and Rosel 2002) and other chromatic anomalies, melanism is the result of genetic mutation(s) (Lamoreux et al. 2010; van Grouw 2013, 2017; Bajpai et al. 2023; Cotts et al. 2023) but is thought to occur much less frequently than does albinism (Sage 1963).



Isabellinism has been infrequently reported among Mammalia but appears to be more common among Aves. This chromatic aberration also is the result of a genetic mutation (van Grouw 2006; Jung and Slough 2012; Jung and Jantunen 2015) and results in a substantial, but incomplete, reduction in melanin. Isabellinism manifests as normally dark pelage appearing as sandy or cream-colored, or as individuals in which coloration appears uniformly “washed out” (Schorger 1973; Everitt and Miskelly 2003; van Grouw 2006; Jung and Slough 2012; Jung and Jantunen 2015). My purpose here is to report the apparent occurrence of isabelline coloration in bighorn sheep, and I provide numerous examples consistent with my contention that isabellinism is a chromatic aberration not previously recognized in that taxon.
Methods
I have maintained observational records of chromatic aberrations in bighorn sheep for nearly 5 decades. During that period, I have discussed anomalous coloration with numerous individuals affiliated with state or federal wildlife agencies, academicians, other individuals familiar with bighorn sheep, and with members of the general public. I asked them to describe any observations of unusual coloration they have seen and recorded those reports and the locations of the observations. I also relied on current bibliographic databases (EBSCOhost, JSTOR, Google Scholar, Web of Science) and a web-based search-engine (Google) to search for words or phrases (albinistic, leucistic, piebald, melanistic, pelage color, etc.) or images specific to abnormal coloration in bighorn sheep and other mammals (Bleich 2017a, 2024). Results were obtained from popular or professional literature, information or photographs archived in electronic databases, and the photographs, electronic images, and detailed descriptions provided by some interviewees. Although many such reports had been published earlier, it remains noteworthy that neither Shackleton (1985) nor McCardle (2012) referred to abnormal coloration in bighorn sheep.
As a result of these explorations, I became aware of subtleties associated with pelage coloration with which I previously had not been familiar, and that indicated several bighorn sheep previously described as being “white” in color were neither albinistic nor leucistic but, instead, were of isabelline coloration. I further used web-based resources to scrutinize several thousand images of bighorn sheep, participated in more detailed discussions with reporting parties, received additional images from colleagues, and delved more deeply into the literature on abnormal coloration (Bleich 2024).
Results
Several decades ago, I learned that “white” desert bighorn sheep occurred in the Clark Mountain Range, San Bernardino Co., California, and a number of years later, J. D. Wehausen informed me of a “white” female desert bighorn sheep he had observed in the Cottonwood Mountains, Riverside Co., California (Bleich 2017a). At the time, Wehausen referred to that individual as being of extremely light coloration, and when redescribing that observation he wrote, “I do not recall details of that animal other than that it looked white compared with the sheep around it” (J. D. Wehausen, Sierra Nevada Bighorn Sheep Foundation, personal communication 21 September 2023). Similarly, W. B. Sloan (U.S. National Park Service, personal communication 21 April 2022) observed a 5-year-old male in the Mesquite Mountains (proximate to the Clark mountain Range; Bleich 2017b), San Bernardino County, California. As described by Sloan, “It’s [sic] white rump patch was almost indistinguishable from the body pelage. Unfortunately, I do not have any quality photos as it was feeding about 800’ away…”. Sloan (personal communication 17 April 2022) also noted, “… the leucistic (?) ram i [sic] saw in the Mesquites in March was the whitest sheep i’ve [sic] ever seen, with it’s [sic] white rump patch being almost indistinguishable from the body pelage. It was much more “white” than several i [sic] see at JOTR [Joshua Tree National Park] that are very light tan in color but still noticeably have the rump patch visible.”
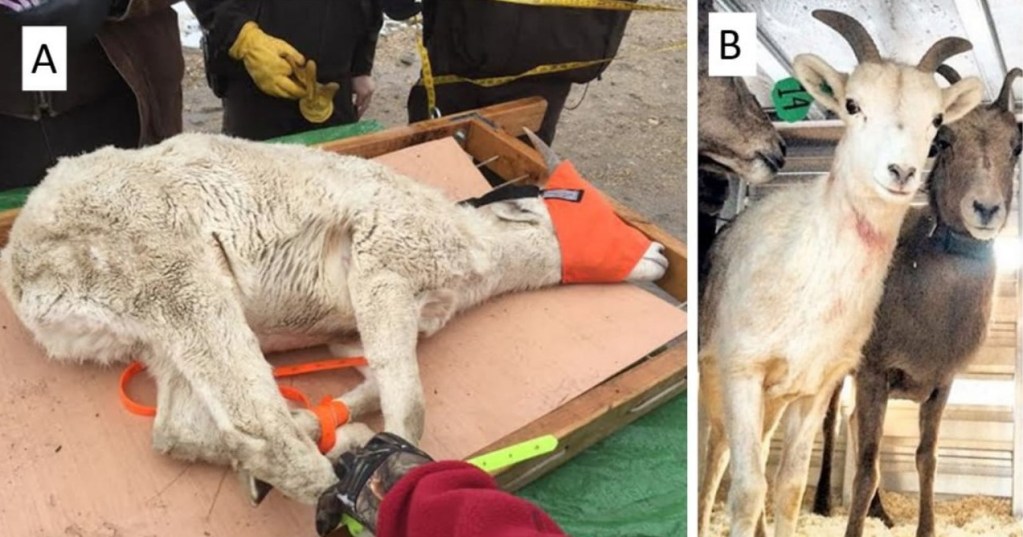
At the time, I did not consider Wehausen’s observation to represent anything beyond normal variation in pelage color, and my initial reaction to Sloan’s report was similar. Having become familiar with additional color aberrations since then (Bleich 2024), however, the details provided by Sloan heightened my curiosity. Moreover, I recently received photographs of apparently isabellinistic bighorn sheep from two additional colleagues. On 7 October 2014, C. Gallinger (Society for the Conservation of Bighorn Sheep) photographed what appears to be an isabellinistic desert bighorn sheep in the Newberry Mountains, San Bernardino Co., California (Fig. 4). That individual, a young male estimated to be 2 years-of-age, was described as not being, “a complete albino as the fur has some color and if you look at the rear hoof it only has a streak of white [and] it is the only sheep I have ever seen that has white horns…” (Gallinger 2023). Further, B. P. Wiedmann (North Dakota Game and Fish Department) observed a mature male Rocky Mountain bighorn sheep (O. c. canadensis) in Dunn Co., North Dakota during the summer of 2022 and described it as being “white” in color. That animal (Fig. 5) is not white but exhibits coloration consistent with the ‘washed-out’ appearance of sandy, or grayish-yellow pelage that normally would be dark and is characteristic of isabellinism, but the rump patch clearly is visible. Subsequently, Wiedmann (personal communication 29 November 2023) reported two additional “white” female bighorn sheep in the same metapopulation occupied by the aforementioned “white” male.


In an effort to further explore the question of isabelline coloration in bighorn sheep, many hours of intensive internet searches yielded at least five additional images of bighorn sheep described as “white” by the photographers (Allynn 2023; Hamilton and Hamilton 2023; Blandford 2023a,b,d), all of which exhibit the uniformly “washed-out” sandy or cream coloration of pelage that normally would be dark. Moreover, each of these animals possess dark eyes and a dark nose, both of which indicate it was not albinistic, and the pelage of leucistic individuals is white (Abreu et al. 2013) and not cream-colored. Perhaps the most striking example of isabelline coloration, however, was revealed upon inspection of an individual described by a contributor as a leucistic young male, and was categorized as such, albeit erroneously, by Bleich (2017a). I recently had the opportunity to view images of that individual, and upon examination it became evident that it was not leucistic but was, instead, of isabelline coloration (Fig. 6).
Discussion
The nomenclature for aberrant coloration has been described as being “muddy,” and neither confined to birds nor to North America by Davis (2007), who noted further that, “… examples of conflicting terminology [exist] in the study of all other vertebrate groups and in other parts of the world as well.” These admonitions were attributed by Davis (2007) to (1) mammals having fewer pigment types than do birds, but that many terms applied to birds were originally proposed for mammals; (2) many descriptive terms having come into use before the mechanisms promulgating color aberrations were understood; (3) color abnormalities occurring unpredictably in the wild and, as a result, being difficult to study; and (4) the misuse of terminology, or misinformation in general. An additional consideration is related to how “normal” coloration (i.e., that absent a genetic mutation; Bernardi et al. 2019) varies, and how it is influenced by polymorphism, geographic variation, or age, wear, or lighting conditions (Davis 2007). Season or stage of molt further add to the potential for unclear determinations among desert bighorn sheep (Nelson 1932), with the result that any assessment of coloration may be somewhat arbitrary (Davis 2007).
When the observations and photographs included or referenced herein (Allynn 2023; Blandford 2023a,b,d; Hamilton and Hamilton 2023; Wildlife Capture Services 2023) are considered along with the misidentifications or confusion of terms, and reports that commonly lack detail, clarity, or consistency, isabellinism is evidenced by the uniformly washed-out appearance of pelage that otherwise would be dark in color, and eyes that are of normal coloration in these images. Bighorn sheep exhibiting this chromatic trait previously have not been described in the literature. As noted by Jung and Jantunen (2015), isabellinism—as indicated in the figures or images referenced herein—may be even more frequent because of the inconsistencies or inaccuracies associated with earlier published descriptions, or as evidenced in photographs available on the internet. Observations of this type are seldomly recorded, and less frequently reported in the literature, but serve to remind others that questions couched in natural history have important implications for the interpretation of adaptation and natural selection (Bleich and Oehler 2000; Bleich 2018) and can provide new insight into earlier conclusions. Indeed, publication of similar observations may encourage others to identify those factors driving the frequency of chromatic aberrations and their associated fitness consequences (Branney et al. 2024).
Isabelline coloration has been reported as a pelage aberration in a number of mammalian orders, and “white” bighorn sheep have been described on numerous occasions across the range of the species (Bleich 2017a). Very light colored (i.e., almost white) bighorn sheep also have been described as “white”, but also may be examples of isabelline coloration given the distinct and clearly discernible white rump patches (Figs. 4–6). Indeed, some of the “white”, “leucistic”, or “albino” bighorn sheep that have been reported (Bleich 2017a, b; this paper) quite likely were isabellinistic individuals that appeared to be white when viewed against a darker substrate, or when seen simultaneously with normally colored individuals (Fig. 5).
The adaptive significance of pelage coloration has been of interest for many decades. Three hypotheses are most apt to explain patterns of color, or coloration itself: concealment, communication, or regulation of physiological processes (Caro 2005). Of these, camouflage (i.e., concealment or crypsis) appears to be the most important evolutionary force explaining overall coloration in mammals, while contrasting patches of pelage evolved primarily for intraspecific signaling (Caro 2005; White 2018; Caro and Mallarino 2022). Whereas natural selection presumably has resulted in benefits associated with coloration or color pattern, alteration of, or a change in, coloration or pattern of color have the potential to affect an individual’s fitness and may be maladaptive (Kaufman 1974; Caro 2005; Dunlop et al. 2019). Alternatively, individual fitness may increase under some circumstances, and eventually lead to a greater representation of a particular color morph in some populations (Kreling 2023). Mutation itself is weak as an evolutionary force (Kreling 2023), however, unless it confers a lethal trait. The rarity of unusual color morphs among bighorn sheep is consistent with the absence of strong directional selection or genetic drift that could result in higher frequencies of a chromatic trait, but two exceptions to this generality may exist. The first is a metapopulation of O. c. nelsoni occurring in the Mojave Desert of California and Nevada (Bleich 2017a, 2017b); the other is in the badlands of western North Dakota, where a number of individual O. c. canadensis previously have been described as being “white” (Fig. 5; Bleich 2017a; B. P. Wiedmann [personal communication 29 November 2023]).
Factors leading to chromatic disorders among wildlife are not well known (Gustalla et al. 2021). Regardless of cause, there likely are fitness costs associated with aberrant, abnormal, or unusual coloration or anomalous color patterns (Caro 2005; White 2018 Dunlop et al. 2019; but see Kreling 2023). For example, rump patches evolved independently in the Artiodactyla (currently Cetartiodactyla; but see Prothero et al. 2022), Lagomorpha, and Rodentia (Guthrie 1971), and presumably conferred some fitness advantage to individuals. Ungulates of the families Bovidae, Cervidae, and Antilocapridae exhibit a rump patch of a color that contrasts strikingly with the individual’s overall body coloration (Bowyer et al. 1991 for review), and sexual dimorphism in rump patches is uncommon (Guthrie 1971). Given the function of rump patches as a potential warning mechanism, signal of group cohesion, or a means of confusing or misleading an attacking predator (Caro 2005; Hogan et al. 2016; Caro et al. 2020), a leucistic, albinistic, or “off-white” (i.e., isabelline) individual may not be effective at intraspecific signaling or evading a predator. Additionally, aberrations in dermal pigmentation or pelage color may confer a lower probability of individual survival because negative pleiotropic effects often accompany chromatic aberrations (Searle 1968; Gratten et al. 2008); among these are impaired visual acuity (Pruthi 2022), reduced auditory function (Lechner and Ladich 2011), or coat color and the associated potential for a negative correlation between body size and fitness, as described by Gratten et al. (2008). Moreover, coat color also serves as an adaptation to environmental conditions and avoiding predators (Kalds et al. 2022). These conditions could decrease an individual’s ability to detect danger, evade danger, or increase the probability of accidental death or injury, and would have concomitant implications for survival and, ultimately, fitness (Wishart 1978; Clark and Jessup 1992; Norrix et al. 1995; Logan and Sweanor 2001; Bleich et al. 2015).
Perhaps most importantly, such abnormally colored individuals are more apt to be noticed and thereby are more susceptible to predation when compared to conspecifics in the same social group (Curio 1976). Thus, there likely exists strong negative selection against bighorn sheep that appear differently than nearby conspecifics, as evidenced by aerial predators that are apt to focus on smaller (i.e., young) individuals (Kennedy 1948; Nette et al. 1984; Bleich et al. 2004) or by terrestrial predators that may be especially cognizant of any animal, juvenile or adult, that somehow appears compromised or otherwise attracts special attention (Jonez 1961; Schaefer et al. 2000; Bleich et al. 2004).
Management Implications
Genomics and its relevance to color variation in some members of the genus Ovis is becoming apparent to investigators (Beraldi et al. 2006; Fariello et al. 2014; Kalds et al. 2022). Recently, chromosome structural variants, epistatic gene interactions, and gene expression have generated much interest among scientists working with domestic breeds (Norris and Whan 2008; Kalds et al. 2022), and interest in those topics is increasing with respect to the vulnerability of North American wild sheep to respiratory disease (e.g., Bowen et al. 2022). Although not considered previously, pelage coloration may in the future become relevant in the context of responses to sources of environmental perturbation such as that described by Duffy et al. (2009), a warming climate (Maloney et al. 2009, 2010; but see Gratten et al. 2010) or, conceivably, be linked to immunological responses and vulnerability to respiratory disease (Ducrest et al. 2008). I suggest investigators record and compile observations of abnormal coloration in bighorn sheep—and perhaps other large mammals (Chen et al. 2019)—if there is a possibility that information of value might be forthcoming as a result of such observations. Doing so will, however, necessitate a more detailed understanding of terminologies, more thorough efforts to distinguish among chromatic aberrations, and a desire to make that information widely available to investigators.
Historically, the majority of aberrantly colored bighorn sheep—and other large mammals for that matter—have been described as albinistic, leucistic, or simply as “white”, but in the absence of detailed scrutiny. If such observations are to have utility in the future, it is essential that investigators are fully aware of the differences in terminologies, the various anatomical parts (i.e., pelage, nose, horns, rump patch, etc.) affected by these chromatic aberrations, and the subtle differences that exist among isabellinistic, leucistic, and albinistic individuals. Minor distinctions are difficult to ascertain in field settings (Holl and Bleich 1983; Bleich 1998), but also when aberrantly colored individuals are encountered during aerial surveys (Bleich 2017a), or through the use of remote cameras, upon which more and more investigators rely, yet may not have the resolution to detect subtle differences (Parker et al. 2020). Moreover, reliability of classifications has been shown to increase with the number of discriminating characteristics (Smith 1988). To that end, I have included a summary of characteristics of which observers must be cognizant if future reports are to be potentially relevant (Appendix I (PDF)). Careful observations and detailed notes based on those observations may enhance our understanding of how bighorn sheep might adapt to the various challenges with which they are faced, or why some populations perform differently than others when faced with similar challenges. In the absence of information that could have been accurately recorded with little extra effort, potentially breakthrough relationships may be missed.
Acknowledgments
I thank R. Weaver (California Department of Fish and Game) and R. Campbell (Society for the Conservation of Bighorn Sheep) who, nearly 50 years ago, called to my attention the ‘white’ bighorn sheep they had observed in the Clark Mountain Range. I also thank J. Wehausen, C. Gallinger, W. Sloan, and B. Wiedmann for sharing details of their observations, and the many agency biologists, guides or outfitters, hunters, photographers, and otherwise interested individuals that participated in detailed discussions or responded to queries and provided useful information. D. Anderson, C. Gallinger, T. Glenner, G. Kerr, R. Robinson, B. Wiedmann, and Wildlife Capture Services, Inc. provided the photographs included in this paper. Comments and suggestions provided by T. Bowyer (University of Alaska Fairbanks), two anonymous reviewers, and the associate editor yielded numerous improvements to the manuscript. This is Professional Paper 149 from the Eastern Sierra Center for Applied Population Ecology.
Literature Cited
- Abreu, M. S. L., R. I. Machado, F. I. Barbieri, N. S. Freitas, and L. R. Oliveira. 2013. Anomalous colour in neotropical mammals: a review with new records for Didelphis sp. (Didelphidae, Didelphimorphia) and Arctocephalus australis (Otariidae, Carnivora). Brazilian Journal of Biology 73:185–194.
- Allynn, D. 2023. Colorado bighorn ewes. Available from: https://www.flickr.com/photos/dawnallynn/18719019135/ (Accessed: 21 August 2023)
- Baccus, J. T., and J. C. Posey. 1999. Melanism in white-tailed deer in central Texas. Southwestern Naturalist 44:184–192.
- Bajpai, V. K., T. Swigut, J. Mohammed, S. Naqvi, M. Arreola, J. Tycko, T. C. Kim, J. K. Pritchard, M. C. Bassik, and J. Wysocka. 2023. A genome-wide genetic screen uncovers determinants of human pigmentation. Science 381:eade6289.
- Baumgartner, L. L. 1943. Pelage studies of fox squirrels (Sciurus niger rufiventer). American Midland Naturalist 29:588–590.
- Bernardi, L. F. O., X. Prous, M. S. Ribeiro, J. Mascarenhas, S. M. C. Genelhu, M. H. Simoes, and T. Bezerra. 2019. First record of albinism for the doglike bat, Peropteryx kappleri Peters, 1867 (Chiroptera, Emballonuridae). Subterranean Biology 30:33–40.
- Beraldi, D., A. F. McRae, J. Gratten, J. Slate, P. M. Visscher, and J. M. Pemberton. 2006. Development of a linkage map and mapping of phenotypic polymorphisms in a free-living population of Soay sheep (Ovis aries). Genetics 173:1521–1537.
- Blandford, T. 2023a. White desert bighorn ram. Available from: https://www.flickr.com/photos/tomblandford/with/16925505315/ (Accessed 1 August 2023).
- Blandford, T. 2023b. White bighorn sheep. Available from: https://www.flickr.com/photos/tomblandford/7050493543/ (Accessed: 21 August 2023).
- Blandford, T. 2023c. Bighorn ewe and lambs. Available from: https://www.flickr.com/photos/tomblandford/9637331658/ (Accessed: 21 August 2023).
- Blandford, T. 2023d. Bighorn contrast. Available from: https://flickr.com/photos/tomblandford/23360103032/ (Accessed: 21 August 2023).
- Bleich, V. C. 1998. Importance of observer experience in classifying mountain sheep. Wildlife Society Bulletin 26:877–880.
- Bleich, V. C. 2017a. Abnormal coloration in bighorn sheep. Bulletin of the Southern California Academy of Sciences 16:54–59.
- Bleich, V. C. 2017b. Leucism in bighorn sheep (Ovis canadensis), with special reference to the eastern Mojave Desert, California and Nevada, USA. Desert Bighorn Council Transactions 54:31–47.
- Bleich, V. C. 2018. Maintaining momentum for conservation: bighorn sheep as an example. Wildlife Society Bulletin 42:540–546.
- Bleich, V. C.2024. Abnormal coloration in Sciurus carolinensis Gmelin (eastern gray squirrel): albinism, leucism, or isabellinism? Prairie Naturalist 56:N1–N4.
- Bleich, V. C., and M. W. Oehler. 2000. Wildlife education in the United States: thoughts from agency biologists. Wildlife Society Bulletin 28:542–545.
- Bleich, V. C., E. F. Cassirer, L. E. Oldenburg, V. L. Coggins, and D. L. Hunter. 2004. Predation by a golden eagle, Aquila chrysaetos, on a juvenile mountain sheep, Ovis canadensis. California Fish and Game 90:91–93.
- Bleich, V. C., C. K. Johnson, S. G. Torres, J. H. Davis, J. M. Ramsey, J. T. Villepique, and B. J. Gonzales. 2015. Psoroptes infestation and treatment in an isolated population of bighorn sheep (Ovis canadensis). Journal of Zoo and Wildlife Medicine 46:491–497.
- Bowen, L., K. Manlove, A. Roug, S. Waters, N. LaHue, and P. Wolff. 2022. Using transcriptomics to predict and visualize disease status in bighorn sheep (Ovis canadensis). Conservation Physiology 10:coac046. https://doi:10.1093/conphys/coac046
- Bowyer, R. T., J. L. Rachlow, V. Van Ballenberghe, and R. D. Guthrie. 1991. Evolution of a rump patch in Alaskan moose: a hypothesis. Alces 27:12–23.
- Branney, A. B., H. N. Abernathy, L. M. Conner, E. Garrison, and M. J. Cherry. 2024. Photographic documentation of melanism in bobcats (Lynx rufus) in the Greater Everglades. Ecology and Evolution 14:e10754. https://doi.org/10.1002/ece3.10754
- Bunch, T. D. and T. C. Nguyen. 1982. Blood group comparisons between European mouflon sheep and North American desert bighorn sheep. Journal of Heredity 73:112–114.
- Caro, T. 2005. The adaptive significance of coloration in mammals. BioScience 55(2):125–136.
- Caro, T., and R. Mallarino. 2022. Coloration in mammals. Trends in Ecology and Evolution 4:357–366.
- Caro, T., H. Raees, and T. Stankowich. 2020. Flash behavior in mammals? Behavioral Ecology and Sociobiology 74:44.
- Chen, L., Q. Qiu, Y. Jiang, K. Wang, Z. Lin, Z. Li, F. Bibi, and 44 additional authors. 2019. Large-scale ruminant genome sequencing provides insights into their evolution and distinct traits. Science 364(6446):eaav620221.
- Clark, R. K., and D. A. Jessup. 1992. The health of mountain sheep in the San Andres Mountains, New Mexico. Desert Bighorn Council Transactions 36:30–35.
- Coetzer, W. G., and J. P. Grobler. 2019. Genetic variation among different springbok (Antidorcas marsupialis) colour variants. Mammalian Biology 99:42–53.
- Conklin Foundation. 2022. The Conklin Foundation. Wild Sheep, Fall 2022:83.
- Cotts, L., J. P. Slifkin, R. Moratelli, L. Goncalves, and O. Rocha-Barbosa. 2023. Multiple colors in anteaters: review and description of chromatic disorders in tamandua (Xenarthra: Pilosa) with reports of new and rare coat colorations. Zoologia (Curitiba) 40:e22034.
- Curio, E. 1976. The ethology of predation. Springer-Verlag, New York, NY, USA.
- Davis, J. N. 2007. Color abnormalities in birds: a proposed nomenclature for birders. Birding 39:36–46.
- Dawson, T. J., and S. K. Maloney. 2008. The significance of fur characteristics in the thermal challenges posed by solar radiation: an underestimated role and source of adaptability. Pages 361–374 in S. Morris and A. Vosloo, editors. Proceedings of the CPB Meeting in Africa: Mara 2008. Molecules to Migration: the Pressures of Life. Medimond International Proceedings, Bologna, Italy.
- Ducrest, A., L. Keller, and A. Roulin. 2008. Pleiotropy in the melanocortin system, coloration and behavioural syndromes. Trends in Ecology and Evolution 23:502–510.
- Duffy, L. K., M. W. Oehler, Sr., R. T. Bowyer, and V. C. Bleich. 2009. Mountain sheep: an environmental epidemiological survey of variation in metal exposure and physiological biomarkers following mine development. American Journal of Environmental Sciences 5:296–303.
- Dunlop, J., D. Peacock, H. Moore, and M. Cowan. 2019. Albinism in Dasyurus species—a collation of historical and modern records. Australian Mammalogy 42:114–118.
- Everitt, D. A., and C. M. Miskelly. 2003. A review of isabellinism in penguins. Notornis 50:43–51.
- Fariello, M. I., B. Servin, G. Tosser-Klopp, R. Rupp, C. Moreno, International Sheep Genomics Consortium, M. San Cristobal, and S. Boitard. 2014. Selection signatures in worldwide sheep populations. PLoS ONE 9(8):e103813.
- Fenson, B. 2017. Genetic freaks: albino, piebald deer and the debate about killing them. Available from: https://www.deeranddeerhunting.com/content/articles/deer-news/genetic-freaks-albino-piebald-deer-and-the-debate-about-killing-them (Accessed: 31 December 2023)
- Fertl, D., and P. Rosel. 2002. Albinism. Pages 16–18 in W. F. Perrin, B. Wursig, and J. G. M. Thewissen, editors. Encyclopedia of Marine Mammals. Academic Press, San Diego, CA, USA.
- Ferris, L. 2013. Wyoming albino bighorn sheep: a trophy among trophies. Casper Star Tribune, 12 December 2013. Casper, WY, USA. Available from: https://trib.com/outdoors/wyoming-albino-bighorn-sheep-a-trophy-among-trophies/article_d6de6fb3-3255-5714-aca2-65ae1af7894d.html (Accessed: 12 December 2013)
- Flores-Peredo, R., B. Bolivar-Cimel, and I. Mella-Mendez. 2021. Albinism in a raccoon (Procyon lotor) from Mexico. Mammalogy Notes 7(2):236. https://doi.org/10.47603/mano.v7n2.236
- Gallinger, C. 2023. White sheep. Available from: https://thewayofthings.org/gallery/white-sheep (Accessed: 5 May 2023)
- Gratten, J., A. J. Wilson, A. F. McRae, D. Beraldi, P. M. Visscher, J. M. Pemberton, and J. Slate. 2008. A localized negative genetic correlation constrains microevolution of coat color in wild sheep. Science 319:318–320.
- Gratten J., A. J. Wilson, A. F. McRae, D. Beraldi, P. M. Visscher, J. M. Pemberton, and J. Slate. 2010. No evidence for warming climate theory of coat colour change in Soay sheep: a comment on Maloney et al. Biology Letters 6:678–679.
- Gustalla, M. G., F. C. de Azevedo, A. Bogliotti, W. F. Lacerda, G. C. Magina, C. A. C. Cardoso, W. Endo, C. A. Peres, T. Haugaasen, D. G. Rocha, and F. G. Lemos. 2021. The mysterious white deer: anomalous coloring in different neotropical deer. Mammalian Biology 101:665–673.
- Guthrie, R. D. 1971. A new theory of mammalian rump patch evolution. Behaviour 38:132–145.
- Hamilton, P., and C. Hamilton. 2023. Rare white bighorn sheep is doing well. Available from: https://www.youtube.com/watch?app=desktop&v=rMrpJU6VCgE (Accessed 11 August 2023)
- Harrison, C. J. O. 1963. Non-melanic, carotenistic and allied variant plumages in birds. Bulletin of the British Ornithologists’ Club 83:90–96.
- Hedges, L., W. Y. Lam, A. Campos-Arceiz, D. M. Rayan, W. F. Laurance, C. J. Latham, S. S. Gopalasamy, and R. Clements. 2015. Melanistic leopards reveal their spots: infrared camera traps provide a population density estimate of leopards in Malaysia. Journal of Wildlife Management 79:846–853.
- Holl, S. A., and V. C. Bleich. 1983. San Gabriel mountain sheep: biological and management considerations. USDA Forest Service, San Bernardino National Forest, San Bernardino, CA, USA.
- Hinton, J. W., K. M. West, D. J. Sullvan, J. L. Frair, and M. J. Chamberlain. 2022. The natural history and ecology of melanism in red wolf and coyote populations of the southeastern United States—evidence for Gloger’s rule. BMC Zoology 7:33. https://doi.org/10.1186/s40850-022-00138-5
- Hoekstra, H. E. 2006. Genetics, development and evolution of adaptive pigmentation in vertebrates. Heredity 97:222–234.
- Hogan, B. G., I. C. Cuthill, and N. E. Scott-Samuel. 2016. Dazzle camouflage, target tracking, and the confusion effect. Behavioral Ecology 27:1547–1551.
- Jacques, C. N., K. R. McCaffery, J. Jenks, and J. T. Baccus. 2008. Semi-melanistic white-tailed deer in northern Wisconsin. The Prairie Naturalist 40:125–128.
- Jones, S. V. H. 1920. Color variations in wild animals. Journal of Mammalogy 4:172–177.
- Jonez, A. 1961. Hunting results in Nevada, 1960. Desert Bighorn Council Transactions 5:87–89.
- Jung, T. S., and J. Jantunen. 2015. Unusual coloration of a North American porcupine (Erethizon dorsatum). Northwestern Naturalist 96:99–100.
- Jung, T. S., and B. G. Slough. 2012. Isabelline coloration in American red squirrels (Tamiasciurus hudsonicus) from the Yukon. Northwestern Naturalist 93:162–164.
- Kalds, P., S. Zhou, Y. Gao, B. Cai, S. Huang, Y. Chen, and X. Wang. 2022. Genetics of the phenotypic evolution in sheep: a molecular look at diversity‑driving genes. Genetics Selection Evolution 54:61. https://doi.org/10.1186/s12711-022-00753-3
- Kaufman, D. W. 1974. Adaptive coloration in Peromyscus polionotus: experimental selection by owls. Journal of Mammalogy 55:271–283.
- Kennedy, C. A. 1948. Golden eagle kills bighorn lamb. Journal of Mammalogy 29:68–69.
- Kreling, S. E. S. 2023. So overt it’s covert: wildlife coloration in the city. BioScience 73:333–346.
- Laacke, R. J., W. F. Laudenslayer, T. Diamond, K. Viotto, and C. A. Long. 2006. Erythrism in the North American badger, Taxidea taxus. Southwestern Naturalist 51:289–291.
- Lamoreux, M. L., V. Delmas, L. Larue, and D. Bennett. 2010. The colors of mice: a model genetic network. John Wiley and Sons Limited, West Sussex, United Kingdom.
- Lechner, W., and F. Ladich. 2011. How do albino fish hear? Journal of Zoology 283:186–192.
- Logan, K. A., and L. L. Sweanor. 2001. Desert puma: evolutionary ecology and conservation of an enduring carnivore. Island Press, Covelo, CA, USA.
- Maloney, S. K., A. Fuller, and D. Mitchell. 2009. Climate change: is the dark Soay sheep endangered? Biology Letters 5:826–829.
- Maloney, S. K., A. Fuller, and D. Mitchell. 2010. A warming climate remains a plausible hypothesis for the decrease in dark Soay sheep. Biology Letters 6:680–681.
- McCardle, H. 2012. Albinism in wild vertebrates. Thesis, Texas State University, San Marcos, TX, USA.
- Morsette, A., Sr. 1996. Hidden man and the two holy men. Pages 184–215 in D. R. Parks, compiler. Myths and traditions of the Arakara Indians. University of Nebraska Press, Lincoln, NE, USA.
- Nelson, E. W. 1932. Mountain sheep. Pages 71–89 in P. N. Gray, editor. Records of North American Big Game. National Collection of Heads and Horns, New York Zoological Society, New York, NY, USA. [Reprint edition. 1990. Derrydale Press, Lyon, MS, USA].
- Nette, T., D. Burles, and M. Hoefs. 1984. Observations of golden eagle, Aquila chrysaetos, predation of Dall sheep, Ovis dalli dalli, lambs. Canadian Field-Naturalist 98:252–254.
- Norris, B. J., and V. A. Whan. 2008. A gene duplication affecting expression of the ovine ASIP gene is responsible for white and black sheep. Genome Research 18:1282–1293.
- Norrix, L. W., D. W. DeYoung, P. R. Krausman, R. C. Etchberger, and T. J. Glattke. 1995. Conductive hearing loss in bighorn sheep. Journal of Wildlife Diseases 31:223–227.
- O’Connor, J. 1974. Sheep and Sheep Hunting. Winchester Press, New York, NY, USA.
- Parker, I. D., R. R. Lopez, and S. L. Locke. 2020. Use of remote cameras in wildlife ecology. Pages 325–333 in N. J. Silvy, editor. The Wildlife Techniques Manual. Eighth edition. Volume I. Johns Hopkins University Press, Baltimore, MD, USA.
- Pirie, T. J., R. L. Thomas, and M. D. E. Fellowes. 2016. Erythristic leopards Panthera pardus in South Africa. Bothalia 46(1):a2034. http://dx.doi.org/10.4102/abc.v46i1.2034
- Prothero, D. R., D. Doming, R. E. Fordyce, S. Foss, C. Janis, S. Lucas, K. L. Marriott, G. Metais, D. Naish, K. Padian, G. Rössner, N. Solounias, M. Spaulding, R. M. Stucky, J. Theodor, and M. Uhen 2022. On the unnecessary and misleading taxon “Cetartiodactyla”. Journal of Mammalian Evolution 29:93–97. https://doi.org/10.1007/s10914-021-09572-7
- Pruthi, S., editor. 2022. Albinism. Mayo Clinic, Rochester, MN, USA. Available from: https://www.mayoclinic.org/diseases-conditions/albinism/symptoms-causes/syc-20369184?p=1 (Accessed: 15 July 2023)
- Sage, B. L. 1963. The incidence of albinism and melanism in British birds. British Birds 56:409–416.
- Sarian, R. 2019. Lopside and the white ewe. California Wild Sheep, Spring 2019:9–10.
- Schaefer, R. J., S. G. Torres, and V. C. Bleich. 2000. Survivorship and cause-specific mortality in sympatric populations of mountain sheep and mule deer. California Fish and Game 86:127–135.
- Schorger, A. W. 1973. The mammals of Dane County. Transactions of the Wisconsin Academy of Science, Arts, and Letters 61:75–85.
- Searle, A. G. 1968. Comparative genetics of coat color in mammals. Logos Press, London, United Kingdom.
- Shackleton, D. M. 1985. Ovis canadensis. Mammalian Species 230:1–9.
- Smith, B. L. 1988. Criteria for determining age and sex of American mountain goats in the field. Journal of Mammalogy 69:395–402.
- Spritz, R. A. 1994. Molecular basis of human piebaldism. Journal of Investigative Dermatology 103:137S–140S.
- Sulem, P. D. F. Gudbjartsson, S. N. Stacey, A. Helgason, T. Rafnar, M. Jakobsdottir, S. Steinberg, S. A. Gudjonsson, A. Palsson, G. Thorleifsson, S. Pa’lsson, B. Sigurgeirsson, K. Thorisdottir, R. Ragnarsson, K. R. Benediktsdottir, J. K. Aben, S. H. Vermeulen, A. M. Goldstein, M. A. Tucker, L. A. Kiemeney, J. H. Olafsson, J. Gulcher, A. Kong, U. Thorsteinsdottir, and K. Stefansson. 2008. Two newly identified genetic determinants of pigmentation in Europeans. Nature Genetics 40:835–837.
- Utah Division of Wildlife Resources (UDWR). 2020. Rare black deer in Moab area died of chronic wasting disease. Utah Division of Wildlife News. Utah Division of Wildlife Resources, Salt Lake City, UT, USA. Available from: https://wildlife.utah.gov/news/utah-wildlife-news/840-black-melanistic-deer-died-chronic-wasting-disease (Accessed: 1 August 2023)
- van Deventer, R., C. Rhode, M. Marx, and R. Roodt-Wilding. 2021. Elucidation of coat colour genetics in blue wildebeest. Mammalian Biology 101:439–449.
- van Grouw, H. 2006. Not every white bird is an albino: sense and nonsense about colour aberrations in birds. Dutch Birding 28:79–89.
- van Grouw, H. 2013. What colour is that bird? The causes and recognition of common colour aberrations in birds. British Birds 106:17–29.
- van Grouw, H. 2017. The dark side of birds: melanism—facts and fiction. Bulletin of the British Ornithologists’ Club 137:12–36.
- van Grouw, H. 2021. What’s in a name? Nomenclature for colour aberrations in birds reviewed. Bulletin of the British Ornithologists’ Club 141:276–299.
- White, T. E. 2018. Cryptic coloration. Pages 1–3 in J. Vonk and T. Shackelford, editors. Encyclopedia of Animal Cognition and Behavior. Springer International Publishing, Cham, Switzerland. https://doi.org/10.1007/978-3-319-47829-6_665-1
- Wildlife Capture Equipment, Inc. 2023. Very rare white bighorn sheep ram lamb caught by HWS. Wildlife Capture Services, Flagstaff, AZ, USA. Available from: https://www.facebook.com/photo?fbid=1914962002062958&set=pb.100054582183543.-2207520000 (Accessed: 24 December 2023)
- Williams, T., S. Kreling, L. Stanton, C. Wilkinson, C. Estien, C. Schell, and E. Carlen. 2023. Of rarity and symbolism: understanding the human perceptions of charismatic color morphs. Research Square Preprints, Durham, NC, USA. Available from: https://www.researchgate.net/publication/373050520_Of_Rarity_and_Symbolism_Understanding_the_Human_Perceptions_of_Charismatic_Color_Morphs (Accessed: 31 August 2023)
- Wilson, L. O. 1970. Whitey, the unique desert bighorn. Desert Bighorn Council Transactions 14:116–122.
- Wishart, W. 1978. Bighorn sheep. Pages 161–171 in J. L. Schmidt and D. L. Gilbert, editors. Big Game of North America: Ecology and Management. Stackpole Books, Harrisburg, PA, USA.