

FULL RESEARCH ARTICLE
David L. Riensche*1 and Meredith Elliot2
1 East Bay Regional Park District, 2950 Peralta Oaks Court, Oakland, CA 94605, USA ![]() https://orcid.org/0009-0002-7255-7919
https://orcid.org/0009-0002-7255-7919
2 Point Blue Conservation Science, 3820 Cypress Drive #11, Petaluma, CA 94954, USA
* Corresponding Author: driensche@ebparks.org
Published 4 Nov 2024 • doi.org/10.51492/cfwj.110.11
Abstract
The Pacific Coast population of the western snowy plover (Anarhynchus nivosus nivosus) is a federally threatened species and is a California Species of Special Concern. Knowledge about their nest site selection requirements is important to this species’ management in the San Francisco Bay area. We measured the percentage of sand, percentage of crushed oyster shells, number of shells, and total shell surface area for 56 nests of western snowy plover at the California least tern (Sternula antillarum browni) colony in Hayward, California, USA, over a 14-year period (2008–2021). Using pairwise t-tests, we compared these measurements to those obtained from 56 randomly chosen non-nest sites. Results indicated that plovers select nest sites with greater oyster shell metrics than paired random sites. The contrast in the shell-related nest metrics (i.e., the difference between the nest site and the paired site) showed significant positive relationships with both hatching success and number of fledglings using linear regression analysis. These results, supporting federal species recovery, suggest that the western snowy plover may improve their nesting success through oyster shell enhancement, which may provide more camouflage for the eggs and chicks than locations with fewer shells.
Key words: Anarhynchus nivosus nivosus, breeding success, oyster shells, San Francisco Bay, threatened species, western snowy plover
| Citation: Riensche, D. L., and M. Elliot. 2024. The importance of oyster shells in the breeding success of western snowy plover). California Fish and Wildlife Journal 110:e11. |
| Editor: Katherine Miller, Wildlife Branch |
| Submitted: 14 February 2024; Accepted: 4 June 2024 |
| Copyright: ©2024, Riensche and Elliot. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Competing Interests: The authors have not declared any competing interests. |
Introduction
One of the rarer shorebirds in North America, numbering around 26,000 birds (Thomas et al. 2012), the snowy plover (Anarhynchus nivosus) is divided into two subspecies (Funk et al. 2007; Page et al. 2009). The western snowy plover (A. n. nivosus) subspecies is a year-round resident along much of the Pacific coastline. The limiting factors for this species include increased predation (Neuman et al. 2004), human disturbance (Ruhlen et al. 2003; Lafferty et al. 2006; Riensche 2021), habitat loss to human development, exotic vegetation, and recreational use (Page and Stenzel 1981; Muir and Colwell 2010). The U.S. Fish and Wildlife Service listed this subspecies as threatened in 1993 (USFWS 1993). The recovery plan approved in 2007 put forward specific goals for safeguarding the long-term conservation of the species by meeting the following: (1) increasing population numbers; (2) conducting intensive, ongoing, and sustainable management for the species and its habitat; and (3) monitoring populations and threats to determine the success of recovery actions and adapting management activities as needed (USFWS 2007).
The western snowy plover is subject to intensive population monitoring (Henkel et al. 2020). Numerous influences can affect plover nest survival including parental conditions (Amat et al. 2001), nest initiation date (Smith and Wilson 2010), and nest location in relationship to the surrounding landscape (Catlin et al. 2011; Colwell et al. 2011, Saalfeld et al. 2011). Diminished nest survival is considered a possible reason for the decline of western snowy plovers (USFWS 2007), with nest failures often caused by mammalian and avian predation, weather (e.g., flooding, hail, and wind), trampling, and human disturbance (Page et al. 2009).
Plovers nest in broad range of habitats and substrates, including: flat, unbroken, barren to sparsely vegetated habitat of coastal beaches, rivers, lake shores, dry pond/lake basins, or large non-vegetated islands (Johnsgard 1981; Bergstrom 1982; Cairns 1982; Cramp and Simmons 1983); dry grasslands (Graul 1975); and alpine and arctic tundra (Cramp and Simmons 1983). Nests are unlined to thinly lined, shallow depressions in hardened clay, silt, loose cobble, pebble, or sand (Johnsgard 1981; Cramp and Simmons 1983). When open-nesting species leave the nest in response to danger posed by a predator, they rely on habitat features to provide a background to hide cryptically patterned eggs. Consequently, the vegetation and substrate characteristics may be habitat features these birds select in a nest site (Colwell 2010). In some habitats, this species selects nest sites with heterogenous substrate consisting of rocks approximately the size of eggs; in finer, sandy habitats, their eggs contrast with the substrate. (Colwell 2010), leading to reduced nest survival (Colwell et al. 2005).
Our goal was to evaluate the effects of oyster shells in breeding success of the western snowy plover within an active California least tern (Sternula antillarum browni) colony in order to aid in management decisions and recovery efforts for this threatened species. Our objectives were (1) to evaluate nest area metrics and (2) to examine the relationships between hatching success and the oyster shell nest area metrics, and number fledged and the oyster shell nest area metrics. We hypothesized that plovers nesting in areas containing higher oyster shell metrics would have improved breeding success. Such data are critical to our understanding of the breeding biology of the western snowy plover and meets the goals set forward in the Western Snowy Plover Recovery Plan (USFWS 2007).
Methods
Study Area
The East Bay Regional Park District manages California least tern, western snowy plover, and black skimmer (Rynchops niger) nesting area at the Hayward Regional Shoreline (37.6297, –122.1461) located along the eastern shore of San Francisco Bay (Riensche 2007; Riensche et al. 2012; Riensche et al. 2015). The San Francisco Bay area is typical of other temperate coastal areas with relative moderate climates not exceeding 35°C. We conducted this study on Island Five (also known as “Tern Town”), a 0.24-ha (0.6-acre) island created from dredge materials. Vegetative cover on Tern Town has been managed (with mechanical techniques and herbicide treatments) to 5–15% over the years to minimize non-native vegetation spread and encourage state and federally listed ground-nesting bird species. A 10 x 20 m grid system was established for nest surveys and outside colony monitoring. The oyster shells were Pacific oyster (Magallana gigas) shells, an introduced species from Asia, which were harvested from Tomales Bay. When the western snowy plover began nesting on Island Five in 2008, no further shell additions or amendments were made.
Data Collection
We located nests by systematically walking through the colony during the breeding season (Riensche 2007). In a 1-m2 area surrounding each nest site, we recorded the following nest area metrics: percentage of area containing crushed oyster shell; percentage of area containing sand; number of oyster shells (> 8-cm2 surface area); shell surface area (in mm2); and average shell surface area (average of surface area [length (in mm) * width (in mm)] measurements of each oyster shell in the 1-m2 of each snowy plover nest site) (Riensche et al. 2015). We measured these same five metrics at a randomly chosen non-nest site within a 5-m radius of each nest site. We then used ArcGIS geoprocessing tools to generate 5-m buffers and random points around known snowy plover nest sites and then generate random, non-nest pair sites within those buffers using the specific geoprocessing tools called “Buffer” and “Create Random Points”. For breeding variables, we collected hatching success (the proportion of eggs that hatched) and number fledged data for each nesting site (Fig. 1).

Data Analysis
We conducted paired t-tests to compare shell-related nest area metrics (percentage of crushed oyster shell, number of oyster shells, total shell surface area, and average shell surface area) of nest sites and paired sites. Using linear regression, we examined the relationships of the two breeding variables (hatching success and number fledged) at each site to the shell-related nest area metrics. In addition, to understand the contrast of the shell metrics at nest sites and paired sites, we used linear regression to examine the relationships between the differences of the shell-related nest area metrics (i.e., [nest site metric] – [paired site metric]) to hatching success and number fledged. We conducted statistical analyses in Stata v16.1 (StataCorp., Stata Statistical Software, College Station, TX, USA).
Results
Over a 14-year period (2008–2021), we monitored a total of 56 nests and recorded nest area metrics (Table 1).
Table 1. Number of nest sites monitored by year.
| Year | # of Nests Monitored |
|---|---|
| 2008 | 1 |
| 2009 | 4 |
| 2010 | 3 |
| 2011 | 3 |
| 2012 | 3 |
| 2013 | 1 |
| 2014 | 3 |
| 2015 | 1 |
| 2016 | 4 |
| 2017 | 9 |
| 2018 | 10 |
| 2019 | 9 |
| 2020 | 3 |
| 2021 | 2 |
| Total | 56 |
Oyster Shell Metrics at Nests and Paired Non-nest Sites
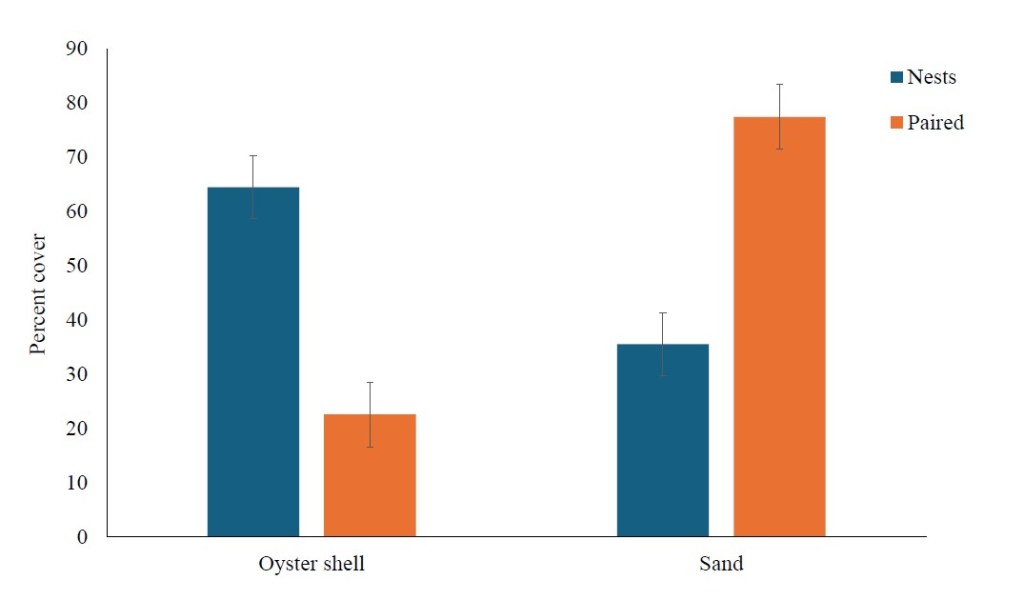
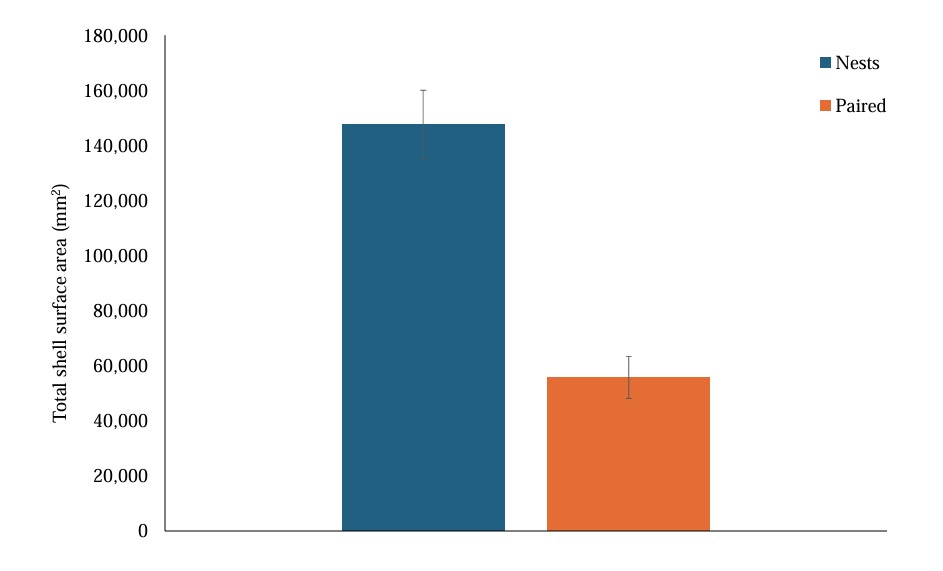
When comparing the shell-related nest area metrics of nest sites and paired sites, we found that average shell surface area was not significantly different between nest sites and paired sites (t = 1.870, df = 55, P = 0.067). Nest sites differed significantly from paired sites for the other three metrics related to oyster shells. Nest sites contained a significantly higher percentage of crushed oyster shell (x̄ = 64.5%) than paired sites (x̄ = 22.6%; t = 9.966, df = 55, P < 0.001; Fig. 2). Nest sites contained more oyster shells with a surface area greater than 8 cm2 (x̄ = 36 shells) compared to paired sites (x̄ = 14 shells; t = 12.227, df=55, P < 0.001). The total surface area covered by shells was also significantly higher at the nest sites (x̄ = 1476.252 mm2) compared to paired sites (x̄ = 557.491 mm2; t = 11.869, df = 55, P < 0.001; Fig. 3).
Relationships between Breeding Variables and Oyster Shell Metrics
When considering the nest area metrics related to oyster shells at the nest site, the only significant relationship between the shell metrics and breeding variables was the number fledged and the number of shells at the nest site (Table 2).
Table 2. Linear regression results between breeding variables and nest area metrics related to oyster shells at the nest site. (*P < 0.05, **P < 0.01, ***P < 0.001)
| Variable | Percent crushed oyster shell | Number of shells | Total shell surface area |
|---|---|---|---|
| Hatching success | B = 0.002, R2 = 0.014, P = 0.387 | B = 0.007, R2 = 0.037, P = 0.156 | B = 1.22×10-6, R2 = 0.017, P = 0.346 |
| Number fledged | B = 0.006, R2 = 0.009, P = 0.479 | B = 0.033, R2 = 0.099, *P = 0.018 | B = 5.74×10-6, R2 = 0.045, P = 0.116 |
The difference in the shell-related nest area metrics revealed positive results in all but seven sites, indicating that most nest sites contained a greater percent area of oyster shell, a greater number of shells, and a larger total shell surface area than the paired site. Linear regression results between the breeding variables and differences in nest area metrics related to oyster shells were significant and positive (Table 3).
Table 3. Linear regression results between breeding variables and differences in nest area metrics at nest sites and paired sites ([nest site metric] – [paired site metric]) related to oyster shells. (*P < 0.05, **P < 0.01, ***P < 0.001)
| Variable | Percent crushed oyster shell | Number of shells | Total shell surface area |
|---|---|---|---|
| Hatching success | B = 0.005, R2 = 0.102, *P = 0.016 | B = 0.011, R2 = 0.100, *P = 0.018 | B < 0.001, R2 = 0.077, *P = 0.036 |
| Number fledged | B = 0.013, R2 = 0.096, *P = 0.020 | B = 0.038, R2 = 0.146, **P = 0.004 | B < 0.001, R2 = 0.098, *P = 0.019 |
Discussion
Worldwide shorebird populations are undergoing startling declines (IWSG 2003; Rosenburg et al. 2019). Of the 57 North American shorebird taxa assessed in 2016, 31 are listed as either high conservation concern, greatest conservation concern, or are listed under the U.S Endangered Species Act (Igelcia and Winn 2021). For shorebirds, many of the effects of climate change have been recognized as threats to their habitats, prey, and natural systems they rely on (Igelcia and Winn 2021). Changes in hydrological cycles (flood/drought), storm intensity and frequency, and coastal habitat losses (flooding and breaching), storm overwash-prone nesting beaches resulting from sea level rise are just some of the challenges climate change creates for shorebirds (Igelcia and Winn 2021; Zeigler et al. 2022). Recent analyses have shown a reduction in winter adult survival with extended cold periods during the winter; with fewer cold snaps expected with climate change, this may lead to reduced adult mortality and might actually help mitigate the many other threats western snowy plovers face (Stenzel et al. 2023). With bird breeding habitat selection involves a series of choices, resulting in a nonrandom selection of territories and nest site. All these choices influence breeding success by affecting the acquisition of resources such as food, water, shelter, mates, nest materials, protection of egg and young from predators (Schoener 1974). Understanding the factors that relate to improved breeding success will help in the recovery of this species.
Our data show that western snowy plovers breed at the Hayward Shoreline nest in areas that contain more oyster shells (in percent coverage, number of oyster shells, and total shell surface area) compared to a randomly selected paired site, although the size of the oyster shells (average shell surface area) was the same. Furthermore, our data show that hatching success and number fledged were greater at nest sites that had more shells (in percent area, numbers, and total surface area) compared to the paired site. These areas of high concentrations of shells may stand out from surrounding areas; this is consistent with other studies where plovers have selected areas with shells for nesting (Page et al. 2009; Tokatlian 2017). Oyster shells are white, presumably reflecting light and keeping the substrate cooler than the surrounding areas of sand; while we do not have temperature data to confirm this, the shells may keep the nesting substrate cooler and improve hatching success. Page et al. (1985) noted alkali flats (which were lighter in color than surrounding sand-gravel substrates) were utilized by nesting plovers more during periods when air temperatures were higher. However, the importance of stationary physical features like oyster shells is difficult to attribute to the number fledged due to the mobile nature of broods.
Nest microhabitat characteristics can potentially affect selection of plover sites (Riensche et al. 2015), survival by altering nest concealment (Colwell et al. 2011), predator detection abilities (Amat and Masero 2004), thermoregulation (Purdue 1976), and the effects of precipitation (Sexson and Farley 2012). The effects of nest microhabitat characteristics on snowy plover nest survival varies among studies and breeding areas (Ellis et al. 2015). Nest site characteristics of a similar species, the piping plover (Charadrius melodus), residing in New Brunswick and Nova Scotia varied with the physiognomy of individual beaches and geographical area (Flemming et al. 1992), which may also be consistent with breeding Snowy Plovers in the San Francisco Bay Region. Additionally, piping plovers nested in sites with more shell cover (Burger 1987).
Predation is the highest cause of nest failure in many shorebird species (Nguyen et al. 2003; Conway et al. 2005; Smith et al. 2007), with predation being the limiting factor in many Charadriidae populations (Johnson and Oring 2002). As ground-nesters, they may mitigate density-dependent nest predation by spacing out their nests (Page et al. 1983), although closer nest distances in plovers was shown to reduce predation risk (Powell 2001). Although we did not quantify predator abundances at our site, the main predators observed included California gull (Larus californicus), common raven (Corvus corax), American crow (Corvus brachythynchos), red fox (Vulpes vulpes), and raccoon (Procyon lotor). We observed predation events by California gull, common raven, American crow, and killdeer (Charadrius vociferus) nesting in close proximity to western snowy plovers (Riensche et al. 2010). Avian predators (e.g., ravens and gulls) begin their search by air for eggs and chicks, and nests that were placed under an object had improved hatching success (Page et al. 1985). While we do not have data on the specific placement of nests, oyster shells, particularly large ones, offer cover to plover chicks at Hayward Shoreline (Fig. 1) and could improve fledging success. In addition, oyster shells may help conceal footprints of plovers that might show up more easily in sand (Powell 2001), thereby reducing detectability by predators and improving hatching success.
The western snowy plover has a serially polygamous, territorial mating system; males and females normally share incubation equally (Colwell 2010), but the males typically care for chicks alone while females pursue additional mates (Page et al. 2009). Consequently, during the long (~ 120 day) breeding season typical of this species along the Pacific coast, females may breed successfully up to three times whereas males may breed twice (Colwell et al. 2007). Their precocial snowy plover chicks are usually out of the nest and foraging independently within 24 hours of hatching. They rely on their parents for thermoregulation and defense and are highly vulnerable to predation until they can fly at 28 to 30 days of age (Page et al. 2009). Survival estimates of color-marked individuals showed higher survival of older (2-year-old) males than females resulting in a population sex ratio biased towards males (Stenzel et al. 2007). Lastly, based on work in coastal northern California by Colwell et al. (2007), roughly 30% of the marked chicks (17% female and 12% male) returned to their natal areas. While none of our plovers were color banded, it appears that the number of breeding birds at this location increased over the 14-year study period, possibly due to some returning to their natal area.
There are many other factors aside from nest characteristics that contribute to the variation we see in hatching success and number fledged of the Hayward Shoreline plovers. Many species of shorebirds depend more on subterfuge and camouflage rather than aggressive antipredator behaviors; therefore, nesting under a “protective umbrella” of some colonial species may reduce predation rates (Burger 1987; Larsen and Moldsvor 1992). Advantages of nesting within a colony include higher detection rates of predators, the ability to deter predators, and “saturation effects” (Siegel-Causey and Hunt 1981; Post and Seals 1993; Emslie et al. 1995). At our study site, the plovers are nesting within an active California least tern colony. It has been shown that piping plover breeding on four coastal beaches in New Jersey, nested closer to eastern least terns (Sterna antillarum antillarum) and had higher reproductive success than those nesting outside tern colonies (Burger 1987). When a predator approached, the terns flew over the predator and dive-bombed them while the plovers walked or flew away, performing distraction displays (Burger 1987). Likewise, in southern California western snowy plovers nest in association with California least terns. Thompson et al. (1997) reported that these terns defend nests and chicks with alarm calls, flights and by diving at and defecating on intruders. Furthermore, in San Diego, California nesting success was greatest for western snowy plovers nesting closest (within 100 m) to an active California least tern nest (Powell 2001), although it was presumed that this benefit was lost after plover eggs hatched. Western snowy plovers at Hayward Shoreline may benefit from these same protections afforded by least terns. As the least tern colony at Hayward Shoreline grew through time (Riensche et al. 2012), so did the number of western snowy plover nests.
There was a large range in breeding effort in this study (2008–2021). Half of the nests in this 14-year study were collected in three years (2017–2019), while another three years only had one nest (2008, 2013, and 2015; Table 1). The higher number of nests in 2017–2019 may be due to nearby areas being flooded out and displaced plovers moving to Tern Town. With such a large range in sample sizes among years, and other factors that contribute to breeding success, the nest site characteristics show a small part of a larger story for this species at Hayward Shoreline. While there was a significant result between number fledged and number of shells at the nest site, we can assume (based on the low R2 value) that the number of shells is not explaining much of the variation in number fledged. Similarly, snowy plovers may be selecting areas with more oyster shells compared to the surrounding landscape, but these significant results also revealed low R2 values, suggesting that the shell variables describe very little about the breeding success of this species at this location. To a small degree, plovers may be actively enhancing their nest site area by moving small shells to their nest site and surrounding area. This species is known to line their nests with shells (Page et al. 2009) and have been observed doing this at Hayward (D. Riensche, pers. obs.). Our results are consistent with other nearby snowy plover breeding sites where oyster shells helped attract plovers to nest, but other factors (e.g., predation) ultimately determined breeding success (Tokatlian 2017). Given the relatively small annual sample sizes and lack of other potentially important covariates (e.g., weather, age of adult plovers, camouflage metrics), future research is needed to understand how oyster shells, amongst other variables, contribute to breeding success. In addition, details on the types of shells, optimal shell density, and alternative materials (which may be more accessible and economical for management) were not examined in this study but could prove useful for future management of this species.
The recovery plan calls for a yearly average production of at least one fledging chick per male being maintained in each of the six recovery units over a five-year period as criteria for delisting (USFWS 2007). Various recovery actions have resulted in growing western snowy plover population with increasing occupancy at breeding and wintering sites throughout their range (Marcot et al. 2021). Management to improve or create nesting habitat for the western snowy plover must anticipate geographical variation in the choice of nest site characteristics and should be designed and implemented based on data from the population of birds under consideration.
The western snowy plover is a heavily managed threatened species, and the effects of such actions on population growth are an immediate concern to conservation biologists. Without intensive mediation, the San Francisco Bay population is likely to decline to an unsustainable level (USFWS 2007). Continuing habitat management for nesting western snowy plovers must reflect the dynamic nature of their nest site selection characteristics. While this species experiences many stressors, our data suggest that oyster shells may provide camouflage for western snowy plover eggs and chicks, thereby improving their breeding success and can enhance our understanding their breeding biology, factors affecting their productivity, and ultimately inform species resiliency and recovery plan goals. Lastly, oyster shell enhancement should be implemented only if local habitat data are available for the birds. Relying on this and other published accounts from other areas may result in inappropriate management strategies.
Acknowledgments
We would like to express our sincere gratitude to the following institutions that made this work possible: U.S. Fish and Wildlife Service, Point Blue Conservation Science, San Francisco Bay Bird Observatory, East Bay Regional Park District, Regional Parks Foundation, U.S. Fish and Wildlife Service Coastal Program, Fremont Bank Foundation, Alameda Countywide Clean Water Community Stewardship Program, New United Motor Manufacturing, Orchard Supply Hardware, Johnston’s and Drake’s Bay Oyster Farms, Las Positas College Science Faculty, California Department of Fish and Wildlife, and California State University East Bay Faculty. Special thanks go to the following volunteers who helped maintain and monitor the site: Patrick Alvarez, George Bloom, Kay Bloom, Emily Brownlee, Nathan Callison, Wade Carlson, Norman Chu, Maggie Clark, Hannah Crosby, Kala Crosby, Rachel Crosby, Janelle Dorcy, Sharon Dulava, Amanda Dwyer, Arthur Garibaldi, Kevin Goe, Tierra Groff, Lisa Henderson, Carin High, Howard High, Kate High, Sam High, Jeanne Jones, Michael Jones, Richard Kaufmann, Anne Krysiak, Sara Lockett, John Mena, Elliot Moon, Martha Morrow, Cody Newell, Caitlin Robinson-Nielsen, Brian Pinomaki, Brianna Pinomaki, Rita Pinomaki, Tyler Pinomaki, Daniel Riensche, Mary Riensche, Nathan Riensche, Rebekah Riensche, Sarah Riensche, Mark Schynert, Adam Shawen, Petra Shawen, Pam Thompson, Pete Thompson, David Wiley, Jon Wiley, and Steve Wiley.
Literature Cited
- Amat, J. A., R. M. Fraga, and G. M. Arroyo. 2001. Variations in body condition and egg characteristics of female Kentish Plover (Charadrius alexandrines). Ardea 89:293–299.
- Amat, J. A., and J. A. Masero. 2004. Predation risk on incubating adults constrains the choice of thermally favorable nest sites in plovers. Animal Behaviour 67:293–300. https://doi.org/10.1016/j.anbehav.2003.06.014
- Bergstrom, P. W. 1982. Ecology of incubation in Wilson’s plover (Charadrius wilsonia). Dissertation, University of Chicago, Chicago, IL, USA.
- Burger, J. 1987. Physical and social determinants of nest-site selection in Piping Plover in New Jersey. The Condor 89:811–818. https://doi.org/10.2307/1368529
- Cairns, W. E. 1982. Biology and behavior of breeding Piping Plovers. The Wilson Bulletin 94:531–545.
- Catlin, D. H., J. D. Fraser, J. H. Felio, and J. B. Cohen. 2011. Piping plover habitat selection and nest success on natural, managed, and engineered sandbars. Journal of Wildlife Management 75:305–310. https://doi.org/10.1002/jwmg.46
- Colwell, M. A., C. B. Millet, J. J. Meyers, J. N. Hall, S. J. Hurley, S. E. McAllister, A. N. Transou, and R. R. LeValley. 2005. Snowy Plover reproductive success in beach and river habitats. Journal of Ornithology 76: 373–382. https://doi.org/10.1648/0273-8570-76.4.373
- Colwell, M. A., S. E. McAllister, C. B. Millet, A. N. Transou, S. M. Mullin, and Z. J. Nelson. 2007. Philopathy and natal dispersal of the Western Snowy Plover. Wilson Journal of Ornithology 119:378–385. https://doi.org/10.1676/06-038.1
- Colwell, M. A. 2010. Shorebird Ecology, Conservation, and Management. University of California Press, Berkeley and Los Angeles, CA, USA.
- Colwell, M. A., J. J. Meyers, M. A. Hardy, S. E. McAllister, A. N. Transou, R. R. Levally, and S. J. Dinsmore. 2011. Western Snowy Plover (Charadrius alexandrinus nivosus) select nest substrates that enhance crypsis and improve nest survival. Ibis 153:303–311. https://doi.org/10.1111/j.1474-919X.2011.01100.x
- Conway, W. C., L. M. Smith, and J. D. Ray. 2005. Shorebird breeding biology in wetlands of the Playa Lakes, Texas, USA. Waterbirds 28:129–138. https://doi.org/10.1675/1524-4695(2005)028[0129:SBBIWO]2.0.CO;2
- Cramp, S., and K. E. L Simmons, editors. 1983. The Birds of the Western Palearctic. Volume 3. Oxford University Press, Oxford, UK.
- Ellis, K. S., J. F. Cavitt, and R. T. Larsen. 2015. Factors influencing Snowy Plover (Charadrius nivosus) at Great Salt Lake, Utah. Waterbirds 38:58–67. https://doi.org/10.1675/063.038.0108
- Emslie, S. D., N. Karnovsky, and W. Trivelpiece. 1995. Avian predation at penguin colonies on King George Island, Antarctica. The Wilson Bulletin 107: 317-327
- Flemming, S. P., R. D. Chiasson, and P. T. Austin-Smith. 1992. Piping plover nest site selection in New Brunswick and Nova Scotia. Journal of Wildlife Management 56:578–583. https://doi.org/10.2307/3808875
- Funk, W. C., T. D. Mullins, and S .M. Haig. 2007. Conservation genetics of snowy plover (Charadrius alexandrinus) in the Western Hemisphere: population genetic structure and delineation of subspecies. Conservation Genetics 8:1287–1309. https://doi.org/10.1007/s10592-006-9278-7
- Graul, W. D. 1975. Breeding biology of the Mountain Plover. The Wilson Bulletin 87:6–31.
- Henkel, L. A., K. K. Neuman, R. W. Stein, and L. E. Stenzel. 2020. Assessing accuracy of sampling schemes to estimate western snowy plover reproductive success. Journal of Fish and Wildlife Management 11:210–216. https://doi.org/10.3996/102019-JFWM-088
- Iglecia, M., and B. Winn. 2021. A shorebird management manual. Manomet, MA, USA.
- International Wader Study Group (IWSG). 2003. Waders are declining worldwide. Conclusions from the 2003 International Waders Study Group Conference, Cadiz, Spain. Wader Study Group Bulletin 101:8–12.
- Johnsgard, P. A. 1981. The Plovers, Sandpipers and Snipes of the World. University of Nebraska Press, Lincoln, Nebraska, USA.
- Johnson, M., and L.W. Oring. 2002. Are nest exclosures an effective tool in plover conservation? Waterbirds 25:184–190. https://doi.org/10.1675/1524-4695(2002)025[0184:ANEAET]2.0.CO;2
- Lafferty, K. D., D. Goodman, and C. P. Sandoval. 2006. Restoration of breeding by snowy plovers following protection from disturbance. Biodiversity and Conservation 15:2217–2230. https://doi.org/10.1007/s10531-004-7180-5
- Larsen, T., and J. Moldsvor. 1992. Antipredator behavior and breeding associations of Bar-tailed Godwits and Whimbrels. The Auk 109:601–608.
- Marcot, B. G., J. E. Lyons, D. C. Elberts, and L. Todd. 2021. Using decision science for monitoring threatened western snowy plovers to inform recovery. Animals 11(2):569. https://doi.org/10.3390/ani11020569
- Muir, J. J., and M. A. Colwell. 2010. Snowy Plovers select open habitats for courtship scrapes and nests. The Condor 112:507–510. https://doi.org/10.1525/cond.2010.090196
- Neuman, K. K., G. W. Page, L. E. Stenzel, J. C. Warriner, and J. S. Warriner. 2004. Effect of mammalian predator management on Snowy Plover breeding success. Waterbirds 27:257–263. https://doi.org/10.1675/1524-4695(2004)027[0257:EOMPMO]2.0.CO;2
- Nguyen, L. P., E. Nol and K. E. Abraham. 2003. Nest success and habitat selection of the Semipalmated Plover on Akimiski Island, Nunavut. The Wilson Bulletin 115:285–291. https://doi.org/10.1676/03-044
- Page, G. W., and L. E. Stenzel. 1981. The breeding status of Snowy Plover in California. Western Birds 12:1–40.
- Page, G. W., L. E. Stenzel, D. W. Winkler, and C. W. Swarth. 1983. Spacing out at Mono Lake: breeding success, nest density, and predation in the Snow Plover. The Auk 100:13–24. https://doi.org/10.1093/auk/100.1.13
- Page, G. W., L. E. Stenzel, and C. A. Ribic. 1985. Nest site selection and clutch predation in the Snowy Plover. The Auk 102:347–353. https://doi.org/10.2307/4086778
- Page, G. W., L. E. Stenzel, J. S. Warriner, J. C. Warriner, and P. W. Paton. 2009. Snowy Plover (Charadrius nivosus). No. 154 in A. Poole, editor. The Birds of North America Online. Cornell Lab of Ornithology, Ithaca, NY, USA. http://bna.birds.cornell.edu/bna/species/154
- Post, W., and C. A. Seals. 1993. Nest associations of Least Bitterns and Boat-tailed Grackles. The Condor 95:139–144. https://doi.org/10.2307/1369395
- Powell, A. N. 2001. Habitat characteristics and nesting success of Snowy Plovers associated with California Least Tern colonies. The Condor 103:785–792. https://doi.org/10.1093/condor/103.4.785
- Purdue, J. R. 1976. Thermal environment of the nest and related parental behavior in Snowy Plovers. The Condor 78:180–185.
- Riensche, D. L. 2007. California Least Tern habitat enhancement and nesting in the East Bay Regional Park District, California, Transactions of the Western Section of the Wildlife Society 43:62–71.
- Riensche, D., C. Robinson-Nilsen, and C. High. 2010. Killdeer observed depredating a Western Snowy Plover nest. Western Birds 41:184–185.
- Riensche, D. L., M. L. Elliott, and S. H. Euing. 2012. Breeding status, nesting densities and diet trends of two endangered California Least Tern colonies. Journal of Environmental Science and Engineering 1:1135–1145.
- Riensche, D. L., S. C. Gidre, N. A. Beadle, and S. K. Riensche. 2015. Western snowy plover nest site selection and oyster shell enhancement. Western Wildlife 2:38–43.
- Riensche, D. L. 2021. Factors influencing the abundance of wintering western snowy plovers at a public beach in California. California Fish and Wildlife Journal, Special Human-Wildlife Interactions Issue:119–129. https://doi.org/10.51492/cfwj.hwisi.8
- Rosenburg, K. V., A. M. Dokter, P. J. Blancher, J. R. Sauer, A. C. Smith, P. A. Smith, J. C. Stanton, A. Panjabi, L. Helft, M. Parr, and P. P. Marra. 2019. Decline of North American Avifauna. Science 366:120–124. https://doi.org/10.1126/science.aaw1313
- Ruhlen, T. D., S. Abbott, L. E. Stenzel, and G. W. Page. 2003. Evidence that human disturbance reduces Snowy Plover chick survival. Journal of Field Ornithology 74:300–304. https://doi.org/10.1648/0273-8570-74.3.300
- Saalfeld, S. T., W. C. Conway, D. A. Haukos, and W. P. Johnson. 2011. Nest success of Snowy Plover (Charadrius nivosus) in the Southern High Plains of Texas. Waterbirds 34: 389–399. https://doi.org/10.1675/063.034.0401
- Schoener, T. W. 1974. Resource partitioning in ecological communities. Science 185:27–38. https://doi.org/10.1126/science.185.4145.27
- Sexson, M. G., and G. H. Farley. 2012. Snowy plover nest survival in Kansas and effective management to counter negative effects of precipitation. Journal of Wildlife Management 76:1587–1596. https://doi.org/10.1002/jwmg.405
- Siegel-Causey, D., and G. L. Hunt Jr. 1981. Colonial defense behavior in Double-crested and Pelagic Cormorants. The Auk 98:522–531.
- Smith, P. A., H. G. Gilchrist, and J. N. M. Smith. 2007. Effects of nest habitat, food, and parental behavior on shorebird nest success. The Condor 109:15–31. https://doi.org/10.1093/condor/109.1.15
- Smith, P. A., and S. Wilson. 2010. Intraseasonal patterns in shorebird nest survival are related to nest age and defense behavior. Oecologia 163:613–624. https://doi.org/10.1007/s00442-010-1644-y
- Stenzel, L. E., G. W. Page, J. C. Warriner, J. S. Warriner, D. E. George, C. R. Eyster, B. A. Ramer, and K. K. Neuman. 2007. Survival and natal dispersal of juvenile Snowy Plovers in central coastal California. The Auk 124:1023–1036. https://doi.org/10.1093/auk/124.3.1023
- Stenzel, L. E., B. R. Hudgens, G. W. Page, K. K. Neuman, A. L. Palkovic, J. L. Erbes, C. R. Eyster, B. A. Ramer, and D. E. George. 2023. Climate change consequences for differential adult survival and the mating system of a temperate breeding shorebird. Ecosphere 14:e4608. https://doi.org/10.1002/ecs2.4608
- Thomas, S. M., J. E. Lyons, B. A. Andres, E. Elliott-Smith, E. Palacios, J. F. Cavitt, J. A. Royle, S. D. Fellows, K. Maty, W. H. Howe, E. Mellink, S. Melvin, and T. Zimmermann. 2012. Population size of Snowy Plovers breeding in North America. Waterbirds 35:1–14. https://doi.org/10.1675/063.035.0101
- Thompson, B. C., J. A. Jackson, J. Burger, L. A. Hill, E. M. Kirsch, and A. W. Atwood. 1997. Least Tern (Sterna antillarum). No. 290 in A. Poole and F. Gill, editors, The Birds of North America. The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologists’ Union, Washington, D.C., USA.
- Tokatlian, K. 2017. Evaluation of oyster shell enhancement on western snowy plover breeding success. Thesis, San Jose State University, San Jose, CA, USA. https://doi.org/10.31979/etd.6tx8-s5y5
- U.S. Fish and Wildlife Service (USFWS). 1993. Endangered and threatened wildlife and plants; determination of threatened status for the Pacific coast population of the western snowy plover. Federal Registrar 58:12864-12874.
- U.S. Fish and Wildlife Service (USFWS). 2007. Recovery plan for the Pacific coast population of the western snowy plover (Charadrius alexandrinus nivosus). Volume 2. California/Nevada Operations Office, Sacramento, CA, USA.
- Zeigler, S. L., B. T. Gutierrez, E. E. Lentz, N. G. Plant, E. J. Sturdivant, and K. S. Doran. 2022. Predicted sea-level rise-driven biogeomorphological changes on Fire Island, New York: implications for people and plovers. Earth’s Future 10:e2021EF002436. https://doi.org/10.1029/2021EF002436