

FULL RESEARCH ARTICLE
Virginia Afentoulis, Andrew Rockriver, Samreen Siddiqui*, and Lauren Damon
California Department of Fish and Wildlife, Bay Delta Region, 2109 Arch Airport Way, Stockton, CA 95206, USA
*Corresponding Author: Samreen.Siddiqui@wildlife.ca.gov
Published 10 July 2023 • doi.org/10.51492/cfwj.109.8
Abstract
The John E. Skinner Delta Fish Protective Facility (Skinner Fish Facility) diverts entrained fish from the State Water Project’s pumping facilities in the southern Sacramento-San Joaquin Delta (the Delta) and transports them via tanker trucks away from the immediate influence of the export pumps. During this process, the fish are counted and released back into the Delta, generally known as fish salvage. Since fish are caught and released back into the Delta, this whole process involves various points that can be stressful to fish, such as the Delta Smelt (Hypomesus transpacificus), a state and federally listed species. To understand this process and improve fish facilities function, an investigation was conducted in 2005–2006, in which we evaluated sublethal stress associated with the terminal portion of the fish salvage process. This study included fish collection, handling, transportation, and release (CHTR) away from the export facilities. Physiological stress parameters for Delta Smelt were measured in the Skinner Fish Facility. We inserted wild and cultured adult Delta Smelt into the CHTR process and later sampled for plasma cortisol, glucose, and lactate, which typically respond to acute and cumulative stress in fish. To help identify the source of overall stress response, we partitioned CHTR into three test phases: collection and handling, trucking and release, and uninterrupted CHTR. Differences in the delayed stress response between the different phases of CHTR were evaluated. Delta Smelt experienced stress in all phases of the CHTR process and in experimental handling controls indicated by higher plasma cortisol concentrations compared to their pre-experiment levels. Wild Delta Smelt experienced higher levels of cortisol response and took longer to recover than did cultured Delta Smelt.
Key words: Contra Costa County, cortisol, fish handling, fish transportation, fish salvage, Hypomesus transpacificus
| Citation: Afentoulis, V., A. Rockriver, S. Siddiqui, and L. Damon. 2023. Delta Smelt stress responses during fish salvage at the John E. Skinner Delta Fish Protective Facility, California. California Fish and Wildlife Journal 109:e8. |
| Editor: Gabriel Singer, Fisheries Branch |
| Submitted: 29 November 2022; Accepted: 13 January 2023 |
| Copyright: ©2023, Afentoulis et al. This is an open access article and is considered public domain. Users have the right to read, download, copy, distribute, print, search, or link to the full texts of articles in this journal, crawl them for indexing, pass them as data to software, or use them for any other lawful purpose, provided the authors and the California Department of Fish and Wildlife are acknowledged. |
| Funding: This project was funded by the CALFED Conveyance Program (CALFED Agreement #4600002115). |
| Competing Interests: The authors have not declared any competing interests. |
Introduction
Delta Smelt (Hypomesus transpacificus) is an endemic osmerid found in the San Francisco Estuary, in California (McAllister 1963; Aasen 1999; Moyle 2002). Delta Smelt populations have been in decline since the mid-1980s and were listed as threatened under both the California Endangered Species Act (CESA) and federal Endangered Species Acts in the early 1990s. In 2009, the CESA status was uplisted to endangered. Since its listing, protection of this species has greatly influenced regional environmental restoration and water management policy. The continued decline in Delta Smelt in mid-2000s triggered court decisions affecting the levels and timing of water exports from the estuary.
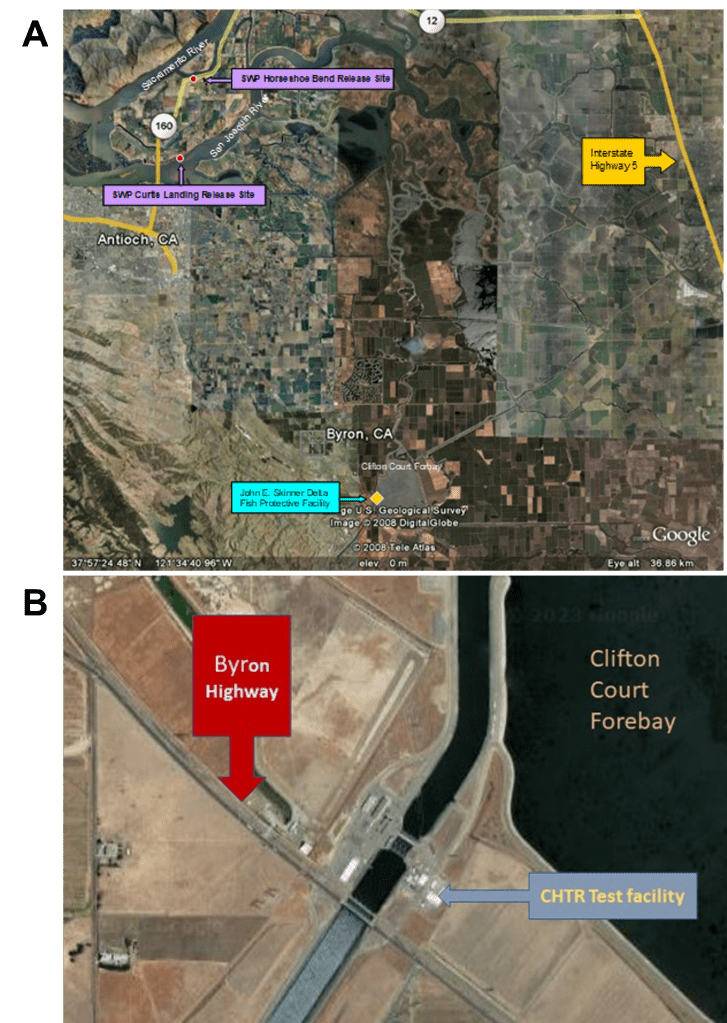
The existing fish collection facilities at the Central Valley Project’s (CVP) Tracy Fish Collection Facility (TFCF) and the State Water Project’s (SWP) John E. Skinner Delta Fish Protective Facility (Skinner Fish Facility) were constructed in the late 1950s and 1960s to protect mainly juvenile Chinook Salmon (Oncorhynchus tshawytscha) and Striped Bass (Morone saxatilis) (Fig. 1). These facilities direct fish away from the intakes of the SWP and CVP pumping facilities by using behavioral dewatering louvers. Louvers concentrate fish, which are funneled and confined into onsite holding tanks for later transport to six release sites in the southern Sacramento-San Joaquin Delta (the Delta), near Antioch, CA, which are considered to be away from the immediate hydrodynamic influences of the export pumps (Morinaka 2013; Skinner 1974). In the late 1990s the California Bay-Delta Program (CALFED) Record of Decision established the goal of reducing entrainment losses associated with SWP and CVP water export facilities through the development and construction of state-of-the-art fish screening (salvage) facilities to assist the restoration of native resident fish species including Delta Smelt. The implementing CALFED agencies were concerned about the feasibility and cost of the proposed south Delta fish screening facilities to protect Delta Smelt.
Although new collection facilities included design improvements to reduce pre-screen predation, increase collection efficiencies, improve hydraulic controls, and provide more fish-friendly holding conditions, the terminal portion of the fish salvage process would remain essentially the same used in the existing facilities. Little was known on the survival or injury of Delta Smelt undergoing the collection, handling, transport, and release (CHTR) phase of the salvage process and there was concern that high mortality in this phase could negate the benefits of improved screening facilities. In addition, Delta Smelt are relatively delicate compared to other species such as those that the fish facilities were designed for (Swanson et al. 2000). Due to these concerns, CALFED requested that the Department of Fish and Game lead a series of collaborative studies (Herrgesell 2012) to assess the acute adverse effects of the existing CHTR process on Delta Smelt and recommend facility improvements and operational procedures to minimize any identified impacts.
We hypothesized that Delta Smelt undergoing the CHTR process would experience high levels of stress due to the degree of crowding and handling typical of these operations. We were concerned about the possibility of adverse impacts since high levels of stress in fish can be immediately debilitating, may decrease the ability of fish to endure subsequent challenges (Schreck 1981, 1982; Barton 2002), and can result in the exhaustion or even death of the animal (Selye 1950). High temperatures, debris, and over-crowding at the fish salvage facilities have, at times, greatly hindered fish salvage and led to high fish mortality (Skinner 1974).
Physiological stress responses in fishes and other vertebrates are grouped as primary (catecholemines and corticosteroids), secondary (systems, i.e., cardiovascular, metabolic, immune, etc.), and tertiary (whole organism) changes at succeeding higher levels of organization (Wedemeyer et al. 1990; Barton 2002). Specific physiological parameters have been employed to assess acute, chronic, and cumulative stress effects at various levels of biological organization over different time scales (Adams 1990). The primary endocrine stress response is marked by elevation of epinephrine and plasma cortisol after perception of a stressful stimulus by the central nervous system (Wedemeyer et al. 1990).
The primary response triggers a secondary cascade response at the blood and tissue level. Epinephrine causes blood glucose to elevate and provide energy for a “fight-or-flight” reaction (Sapolsky et al. 2000). Cortisol helps sustain elevated glucose levels through depletion of tissue glycogen (Shanghavi and Weber 1999). Elevation of lactate is caused by either fright or severe muscular exertion (Wedemeyer et al. 1990; Wendelaar Bonga 1997). Other secondary compensatory stress responses may include lowering of plasma chloride, hemorrhage of the thymus, hypertrophy of the internal body, and immunosuppression (Wedemeyer 1996).
Stress responses are considered adaptive and compensatory if physiological changes allow the organism to maintain homeostasis in the face of biotic or abiotic challenges. However, with prolonged or severe exposure, physiological and whole-animal responses may become detrimental to the health of the organism and be considered maladaptive (Barton 2002). Response cascades to this level can produce effects at the individual and population level (tertiary response) through reductions in growth, infectious disease resistance, reproductive success, and survival (Wendelaar Bonga 1997, Schreck et al., 2001).
The purpose of this investigation was to identify practical and reliable procedures to measure sublethal stress for use in future fish facilities evaluations, were a new facility built. The first objective was to evaluate a set of biological stress indicators and their ability to measure the magnitude of Delta Smelt stress response and recovery time within the CHTR process. The second objective was to identify which components of the CHTR process were the most stressful to Delta Smelt. These results would also act as the first step toward determining likely tertiary stress responses in salvaged Delta Smelt (V. Afentoulis, CDFW, unpublished data). The stress results would be used along with data from the other CHTR studies to help design new salvage facilities or make improvements to existing facilities.
Methods
All experiment replicates were conducted at the Skinner Fish Facility from March through May 2005 and from December 2005 through May 2006. Prior to use, Delta Smelt were held in tanks with treated aqueduct water at the University of California, Davis’s (UCD) Fish Conservation and Culture Laboratory (FCCL). Experiment replicates took place in the holding tank buildings of the Skinner Fish Facility or at the neighboring CHTR test facilities (Fig. 1). The Skinner buildings have 7 large (6.1 m deep x 6.1 m diameter) holding tanks which temporarily collect fish diverted (salvaged) from water heading towards the SWP pumps.
The CHTR phase is only one portion of a series of salvage processes that entrained fish must negotiate successfully to avoid being lost (preyed or entrained in water to be exported) within the SWP. Fish enter the headworks of the SWP export facilities through Clifton Court Forebay (Skinner 1974). Prior to the export pumps, the Skinner Fish Facility uses a set of primary louvers (behavioral barriers) to divert fish into a secondary bypass channel. Fish are then guided from the secondary channel into holding tanks using either louvers or positive barrier fish screens. Fish are continuously collected and held in the holding tanks for a period ranging from 8 to 24 hours based on species (no longer than 8 or 12 hours for Delta Smelt and Chinook Salmon, respectively, and up to 24 hours for all other species based on operational conditions). The CHTR process begins by draining each holding tank and its contents into an 1,893-L loading bucket located in the bottom center of the tank (collection phase). The loading bucket is raised via a crane and moved to a 9,463 or 10,599-L tanker truck, and the contents of the bucket are released (handling phase) through stationary fish release pipes. The tanker truck is driven approximately 50 minutes to one of six release sites in the Delta (transport phase); fish are then released into the Delta (release phase).
Test Fish
We used adult wild and cultured Delta Smelt in our experiment replicates. Wild Delta Smelt were collected in late fall in 2004 and 2005 from the western Delta using modified methods developed by Swanson et al. (1996). We collected fish using a modified lampara net with a landing bag with a stretch mesh size of 0.95 cm. Wild smelt were transported to the FCCL and held from 1–3 months before use in experiments. The FCCL expanded their facility to accommodate the increased demands of test fish and holding facilities for CHTR studies (Baskerville-Bridges et al. 2006).
Both wild and cultured Delta Smelt were held and cared for in a similar manner (Baskerville-Bridges et al. 2005, 2006). Cultured (F1 generation) Delta Smelt were propagated from wild brood stock. The adult cultured smelt were held in black 550-L, circular holding tanks (1.52 m diameter x 61 cm deep) until needed for the CHTR experiment replicates. Delta Smelt were fed a mixture of dry pellet feed (Kyowa 1000-c) and Hikari plankton. Delta Smelt were fed twice daily, up until the day of the experiments. The day of the experiments, fish from a selected tank were not fed until after removal of the test fish, so that test fish would not be run in experiments with full stomachs. After each experiment, Delta Smelt were held in 4 smaller tanks at the CHTR test building in minimally treated water (sand filtered and UV treated) from the Clifton Court Forebay outlet channel. Water temperatures in test building tanks ranged from a low of 9°C in winter to a high of 22°C in spring, depending on the temperature of ambient Delta intake water temperature. We measured water quality parameters including dissolved oxygen (mg/L), water temperature (°C), and electrical conductivity (μS/cm), which we monitored before, during and immediately after each experiment. Water quality was also measured within the storage tubs holding Delta Smelt at the 2-, 24-, and 48-hour post-treatment intervals. Water quality data is available in the appendices and varied seasonally, and with changing conditions.
Physiological Measurements
We collected and processed blood samples using a protocol adapted from Young et al. (2001). Four smelt were netted from a bucket and wrapped in a Kimwipes® tissue before bleeding. Each smelt’s caudal fin was immediately severed with a scalpel just anterior to the caudal peduncle. Blood was collected from the exposed caudal vessels using standard microhematocrit tubes. The amount of whole blood drawn from each individual varied from 2–20 µL. We pooled plasma from individual fish to meet the minimum required volume for cortisol analysis of 5 µL. Once filled with blood, the microhematocrit tube was sealed. Once bled, fish were stunned by a blow to the head and euthanized by sectioning the spine just posterior to the skull with a scalpel. Any Delta Smelt that was mishandled or dropped during any phase of the experiment or blood collection process was excluded. Each fish was measured (fork length), weighed, and then frozen for reference purposes.
Blood samples from the microhematocrit tubes were processed in two ways. We centrifuged blood at 9,500 rpm for 5 minutes to separate packed red cells from plasma using a Vulcon Technologies Microspin 24TM centrifuge. Additionally, approximately 25% of the whole blood samples were immediately measured for either glucose or lactate concentrations. We obtained glucose readings using an Accu-Chek® glucose meter. An AccuSport™ meter was used to measure lactate concentrations. Hematocrit values were obtained by measuring the centrifuged hematocrit tubes on a graduated chart for percent packed blood cells within blood samples. Values obtained were recorded to the nearest 1%.
We transferred blood plasma from the centrifuged tubes into 1.5 mL microcentrifuge freezer vials. Because a minimum of 5 mL of plasma was needed for cortisol analyses, plasma from a minimum of two fish was pooled before storage at –40°C. Frozen plasma was transported on dry ice to the UCD Population Health and Reproduction’s Clinical Endocrinology Laboratory for blood plasma cortisol measurements. We used enzyme-linked immuno-sorbent assay (ELISA) to determine blood plasma cortisol levels (Munro and Stabenfeldt 1985).
Experimental Design
We tested 240 fish in CHTR events for each of the processes or treatments separately: collection and handling (C+H), 21 events; transportation and recovery (T+R), 18 events; full CHTR, 19 events; and net stress (NS), 28 events. For each of the events, 24 randomly selected fish were used for each experiment replicate, with wild and cultured fish tested in separate replicates. Before each replicate (for either wild or cultured replicates), we randomly selected and sampled four fish 30 minutes before the experiment (–0.5-hour time interval) to measure pre-treatment baseline levels. Measurements from these fish were referred to as the negative or pretest control condition, because they experienced no CHTR or NS event. While not a true negative control, the baseline is the reference point and elevation above this undisturbed-state reference is what indicates stress. We placed the remaining 20 fish into the trials.
After trial completion, we immediately sampled four fish (0-hour post-treatment time interval) for blood collection within 2 minutes. Another four fish were placed in a black 19-L bucket for blood collection at the 0.5-hour post-treatment time interval (technique developed for Delta Smelt stress assessment by Swanson et al. 2005). We placed the remaining 12 fish in random groups of four, into three separate 83-L storage tubs (Rubbermaid®) placed in a black 1,136-L holding tank with a flow-through water supply. At 2-, 24-, and 48-hours post-treatment time intervals, a group of four fish were removed for physiological testing. All remaining fish were used for blood collection by the time the 48-hour observations were completed. Once bled, fish were stunned by a blow to the head and euthanized by sectioning the spine just posterior to the skull with a scalpel.
Experimental treatments consisted of testing fish exposed to C+H, T+R, the full CHTR process, and NS. The C+H treatment simulated routine fish handling when fish are transferred from the Skinner Fish Facility holding tanks into the loading bucket and released into a 9,463-L transport truck. Delta Smelt were moved between the pre-treatment holding tanks to the experiments and back to the CHTR test building. Fish were netted before each of the CHTR processes using a soft-mesh brine shrimp net and transferred into a filled 19-L bucket. To start the experiment, we lowered the insertion bucket by hand-line into the holding tank. Once the bottom of the bucket was just below the water surface, it was slowly inverted to empty the fish. The 20 fish were allowed to acclimate in the holding tank for between 5–20 minutes before the holding tank started draining. Once the tank drained and the fish were concentrated into the loading bucket, we raised the bucket and emptied it into an 1,893-L fiberglass retrieval tank. We removed Delta Smelt from the retrieval tank using a brine shrimp net as quickly as possible (in about 5 minutes). Four random fish were immediately sacrificed for physiological testing and the remainder held for 0.5, 2, 24, or 48 hours.
The T+R treatment simulated transport of Delta Smelt in the fish tanker truck from the Skinner Fish Facility to the Delta release locations, and the subsequent release of the tanker’s contents through a pipe into the river. Delta Smelt were loaded into the transport truck using the 19 L bucket inversion technique described in the C+H treatment. Once all the fish were loaded, the tanker truck drove for approximately 50 minutes to approximate the travel time to the Delta release sites. Upon return to the Skinner Fish Facility, the truck drove up a ramp to the top of a simulated release apparatus. Below the ramp was a 45,425-L above-ground pool (8.78 m long x 4.57 m wide x 1.22 m deep). We released the contents of the tanker truck from the back of the truck into the previously filled pool through a large pipe (25.4 cm diameter x 7.62 m long). The simulated release apparatus was shorter in length and lacked some features of the actual release sites due to site limitations. The pool water was allowed to drain through a drum screen at the opposite end of the pool. The water in the pool was then drained to concentrate the fish in a recessed trough (457 cm long x 46 cm wide x 30 cm deep) at the bottom of the pool. The draining took about 15 minutes after which 4 fish were removed using a brine shrimp net and sampled.
The CHTR treatment simulated the entire process of collecting, handling, transporting, and releasing salvaged Delta Smelt. Delta Smelt were inserted into the holding tank and went through the same process as described in the C+H experiment. However, instead of emptying the loading bucket into the retrieval tank, the bucket emptied its contents directly into the tanker truck. The truck then drove around for 50 minutes, returned, and emptied its contents into the recovery pool as described in the T+R experimental design. Delta Smelt were collected and bled in groups of four individuals.
The net stress (NS) was a positive-control treatment, which imitated the stress of netting and transporting Delta Smelt. This was done by netting fish into a 19-L bucket, the same procedure by which all experiment fish were both introduced to and retrieved from all the CHTR treatments. At the Skinner Fish Facility, Delta Smelt were netted and placed in a 19-L bucket. The bucket was transported a short distance to mimic the transport of pre-treatment fish to the holding tank or tanker truck. Instead of releasing smelt into the tank or truck, fish were inserted into a pre-filled black 3,407-L circular tank to simulate the insertion process. Fish were acclimated in the circular tank for approximately 5 minutes and then removed from the tank with nets to simulate handling during the CHTR, T+R, and C+H experiments and subsequent netting to remove them from the experiments. This process lasted approximately 15 minutes. These fish were held and bled identically to the other CHTR event treatment groups for up to 48 hours. The netting from source tanks, confining in a bucket, transfer to another container, and subsequently netting are all known stressors for fish (Carragher and Rees 1994).
Statistical Analyses
A preliminary two-sample t-test analysis comparing 2005 and 2006 data showed no statistical differences; therefore, results of the two study seasons were pooled for statistical analysis. We examined the time-response of the selected parameters by plotting their mean values with their respective standard errors. The variability around means of environmental data was described using their standard deviations. Systat® 11 software was used to perform all statistical analyses.
We tested our data for the suitability for parametric analysis. Normality was tested by means of graphical plots of the data. Kurtosis was evaluated using the Shapiro-Wilk statistic and adjusted through log transformation of the data. Homogeneity of variance was tested using the Bartlett test (Zar 1999). Some data were not normal, but we deemed that our data were suitable for ANOVA and post-hoc testing given the robustness of the statistical tests. Non-parametric tests were not used because the risk of committing a type one error is extremely small (less than a 0.07 probability for sample sizes greater than 10) given the sample sizes and variances (Zar 1999). The data was also log-transformed, and results remained unchanged from the untransformed data.
We compared mean cortisol, glucose, and hematocrit concentrations among treatment types (NS, C+H, T+R and CHTR) for each time interval using ANOVA; Tukey’s multiple comparison test was used to identify statistically significantly differences between means (a = 0.05). To determine if post-treatment physiological parameters differed significantly from control conditions (–0.5-hour control or baseline condition samples), we used ANOVA and post hoc Dunnett’s test to detect differences between pretest control condition levels and each post-treatment time level (0.5, 2, 24 and 48 hours). We used t-tests to detect physiological parameter differences between wild and cultured Delta Smelt at each time interval for each treatment type. Background mortality was taken into account and compared with the same population over time held at the UCD Fish Conservation and Culture lab and were found to be the same, approximately 1%.
Quality Control for Field Measurements
The YSI 556 dissolved oxygen meter was calibrated daily before use by the instrument’s preprogrammed procedures. We calibrated the Acculab scales using a 200 g reference weight. The electrical conductivity sensor of the YSI 556 and the Accu-Chek glucose meter were checked before each study season (the frequency of which is according to manufacturer’s specifications) using commercial standard solutions. The AccuSport lactate meter was calibrated using the manufacturer’s calibration strips before each use. The accuracy of the other field measurements was not determined.
We took precision (repeat) measurements at a minimum frequency of 5% of the total observations recorded for each parameter. An average of 8.9% of the hematocrit, fork length, body weight, electrical conductivity, dissolved oxygen, and water temperature readings were repeated either by the instrument operator (precision) or between two individuals (e.g., hematocrit and fork length). These repeat values were used to determine the percent deviation from the average of the two values. We took percent deviation measurements on field data collected from 17 of the 80 experiments. Less than 5% deviation was the performance criterion for these readings. The precision of the glucose or lactate readings was not determined. Accuracy readings of field measurements taken by the YSI 556 water quality instrument, conductivity, DO, and temperature, were not attempted.
Cortisol Analysis
Plasma samples were analyzed for cortisol levels by the UCD Endocrinology Lab. Cortisol antiserum (R4866) was combined with horseradish peroxidase (HRP) to yield a conjugate used to coat microtiter plates (Immulon 1) where the competitive enzyme reaction occurs. Aliquots of each extracted plasma sample were added to the wells on each plate for competitive action with antiserum-HRP conjugate. After rinsing, a color developer substrate solution was added to determine the amount of conjugate (cortisol-HRP) bound to the plate wells. Percent absorbance at a wavelength of 405 nm was inversely proportional to the amount of bound cortisol.
Absorbance was measured by an automatic microtiter plate spectrophotometer (Molecular Devices Vmax). Instrument data were gathered using an interfaced computer for later analysis. Calibration curves were constructed using weighted least-squares linear regression according to the method of Rodbard and Lewald (1970). We reduced the raw data by logit-log transformation to yield concentration of plasma cortisol in ng/mL.
Laboratory quality assurance checks for the cortisol values consisted of laboratory controls and duplicate measurements of all plasma samples analyzed. ELISA analysis sensitivity was determined as the least amount of hormone that could be distinguished from zero concentration of standard as defined by the mass at 2 standard deviations above the mean of the zero absorbance. Recovery rates of selected samples were determined by analyzing results of serial dilutions compared against a cortisol standard curve. The precision of the ELISA analyses was assessed in selected pooled serum samples by repetitively measuring the same sample and calculating the coefficient of variation (CV).
Results
Number of Tests
A total of 2,088 Delta Smelt were sampled from 86 experiment replicates. The number of completed replicates for each experiment type varied from a low of 6 T+R replicates with cultured Delta Smelt to a high of 13 NS replicates with wild Delta Smelt. A greater proportion of the experiments used wild Delta Smelt. Mortality rates of test Delta Smelt averaged 1.6% overall with most mortalities found at 48 hours post experiment. Pretest fish mortality rates at the culture facility were in the range of 1.3%, therefore mortality was not considered to be a factor related to stressful events during or after experiments.
The number of physiological measurements varied among treatment groups mainly due to varied amount of plasma from each fish and the need to pool plasma samples. Cultured Delta Smelt were relatively larger and heavier than wild Delta Smelt used in our study. Mean fork length for wild Delta Smelt was 64 ± 2.0 mm (n = 1,065) and was 68 ± 2.3 mm (n = 860) for cultured Delta Smelt. Mean wet weight for wild Delta Smelt was 2.1 ± 0.02 g (n = 1,065) and 2.7 ± 0.05 g (n = 862) for cultured Delta Smelt.
Water Quality and Holding Conditions
Water quality and holding conditions during the two experimental seasons were deemed appropriate for conducting our studies and reflected seasonal and operational conditions at the Skinner Fish Facility. Water temperatures ranged from a low of 9°C in winter to a high of 22°C in spring (for both years combined) and averaged 15 ± 3°C. Water temperatures stayed well within critical temperature maxima and minima for this species (Swanson and Cech 1996). Dissolved oxygen levels varied with water temperature from 6 mg/L to as high as 15 mg/L (due to the injection of compressed oxygen during fish transport phase), all acceptable levels for holding and transporting fish (Portz et al. 2006). Dissolved oxygen readings averaged 10 ± 2 mg/L.
Electrical conductivity readings from pre-experimental holding tanks and collection tanks in the Skinner Fish Facility were generally similar. Electrical conductivity for the compared NS and C+H replicates ranged from 96 to 416 µS/cm and averaged 185 ± 63 µS/cm. However, Delta Smelt in the T+R and CHTR experiments experienced higher electrical conductivity in the tanker truck and collection pool due to the standard operating procedure of adding salt (NaCl) to the tanker truck water. The electrical conductivity in the truck tank varied from 2,277 to 9,452 µS/cm and averaged 4,772 ± 1,298 µS/cm which are within ranges appropriate for fish (Portz et al. 2006).
Wild Delta Smelt Responses
Cortisol.—Plasma cortisol of wild Delta Smelt was elevated immediately after each treatment and remained elevated for at least until the 2-hour sampling point (Figs. 2 and 3). For most treatments, except for T+R, cortisol was significantly elevated from control condition level at all post-treatment time intervals. Generally, wild Delta Smelt cortisol did not return to pre-treatment levels prior to 24 hours post-treatment (Fig. 2).
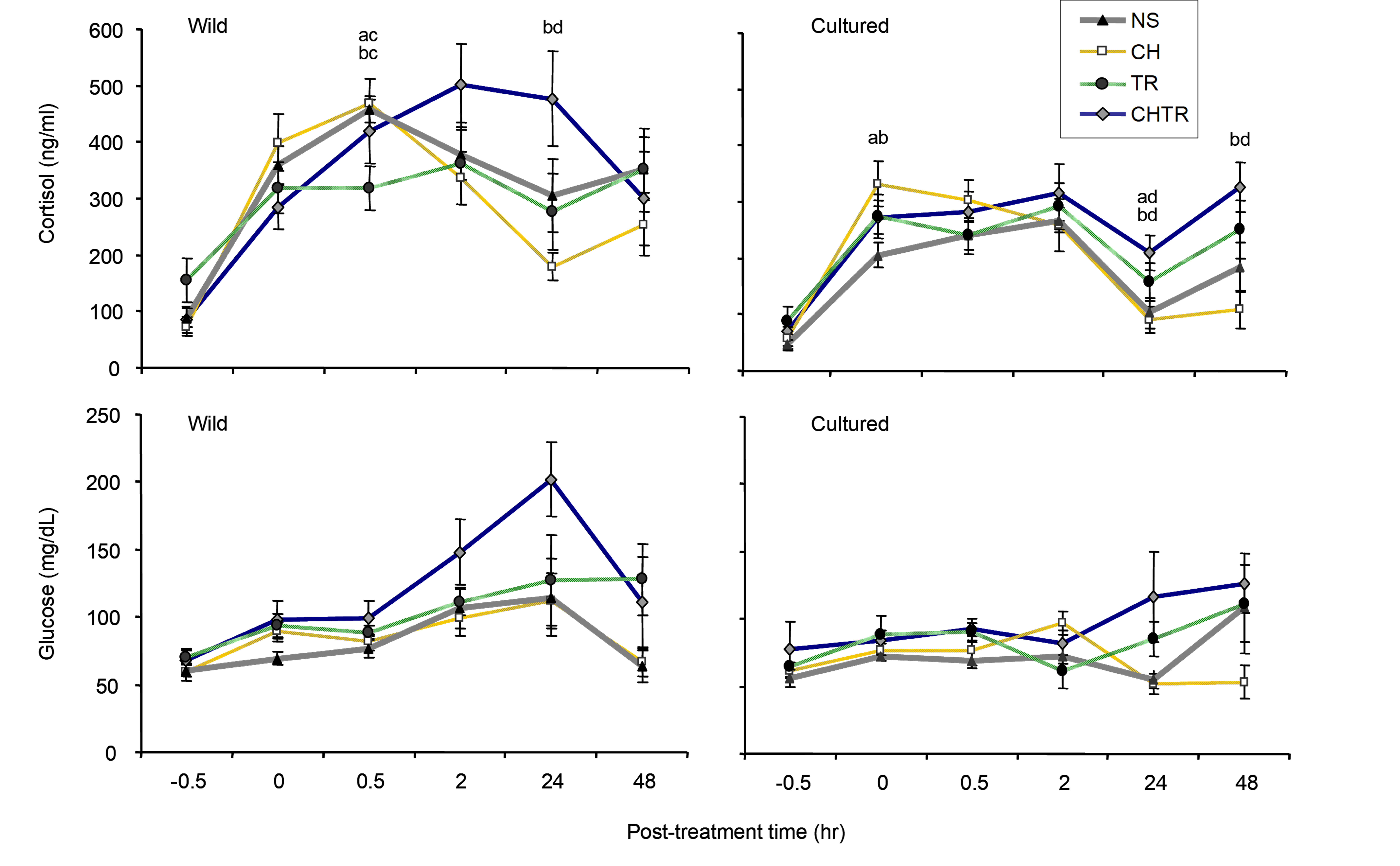

Mean cortisol levels were similar among all post-treatment time intervals when comparing treatments (NS, C+H, T+R, and CHTR). However, significant differences occurred between some of the treatments (Fig. 2). At 0.5-hour post-treatment, wild Delta Smelt in the T+R had significantly lower cortisol levels than wild Delta Smelt in the C+H and NS treatment events. At 24 hours, wild smelt in the CHTR replicates had the highest cortisol levels of any of the treatments. This difference was only significant between the CHTR and the C+H treatments.
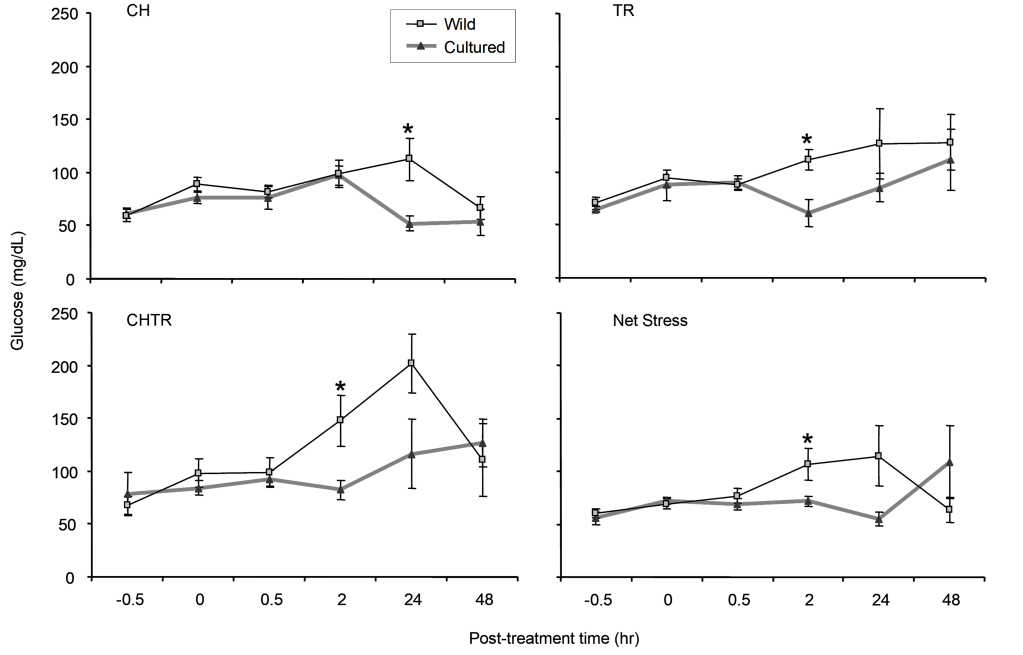
Glucose.—Wild Delta Smelt glucose levels rose moderately from the -0.5 hour (Fig. 2). Glucose levels at 2- and 24- hours post-treatment was significantly different from pre-treatment levels for all three treatments: NS, C+H, and CHTR. Glucose levels for the T+R replicates were similar to -0.5-hour levels for all post-treatment intervals, except for an uptick at 48 hours post treatment that wasn’t significant. By 48 hours, glucose was no longer significantly elevated for each treatment compared to pre-treatment levels (Fig. 2). In addition, there were no significant statistical differences in means among NS, C+H, T+R and CHTR treatments at each sampling period (Fig. 2). The CHTR treatment showed an increase above the other treatments after 2 hours; however, this increase above the other treatments and above –0.5 hours was not statistically significant (Fig. 2, P > 0.05). However significant differences between cultured and wild Delta Smelt were noted at the 2- and 24-hour intervals (Fig. 4, P < 0.05).
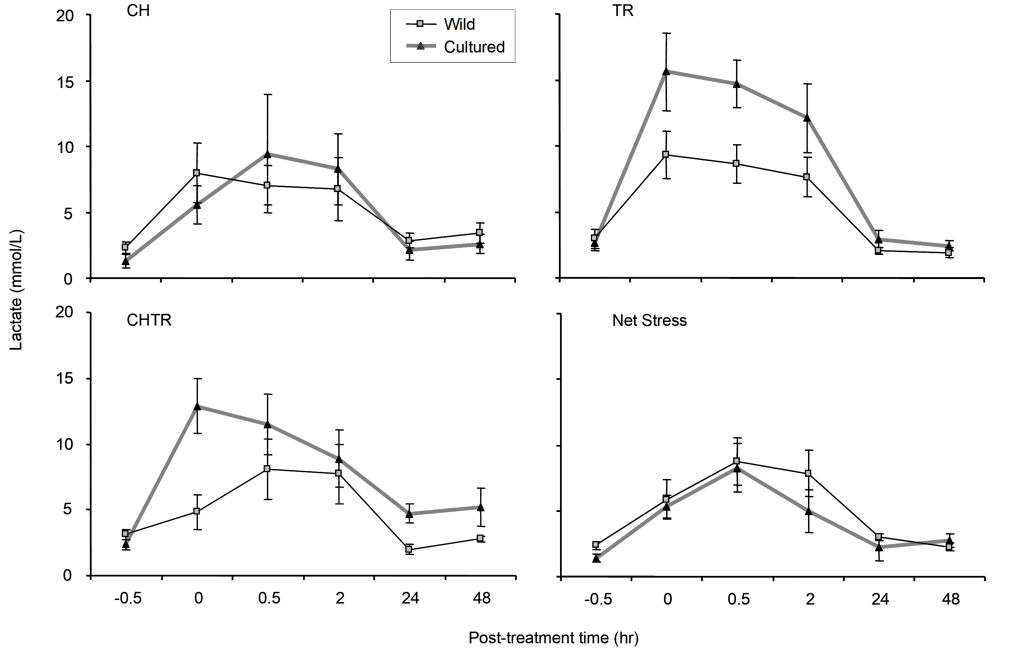
Lactate.—We made no statistical comparisons among tests for lactate because the lactate meter’s automatic shut-off could not be disabled and the required blood volume could not always be met. Since this was a sports medicine device for rapid reading, we also obtained numerous non-numeric readings likely due to programming limitations.308 numeric values were recorded for lactate (Table 1).
Table 1. Number of lactate values by time interval in each experimental treatment: collecting and handling (C+H), transportation and recovery (T+R), full collection, handling, transportation, and release (CHTR), and net stress (NS) during the experiments at the John E. Skinner Delta Fish Protective Facility, Tracy, CA, USA, 2008.
| Time Interval | C+H | T+R | CHTR | NS |
| –0.5 | 10 | 11 | 15 | 11 |
| 0 | 11 | 15 | 16 | 17 |
| 0.5 | 10 | 15 | 13 | 14 |
| 2 | 13 | 13 | 15 | 16 |
| 24 | 9 | 14 | 13 | 14 |
| 48 | 9 | 9 | 10 | 15 |
However, wild Delta Smelt apparently had a range of elevated lactate levels at each post-test time interval and among treatments, with most of the elevation occurring within the first (0-hour) post-treatment interval lasting until the 24-hour interval (Fig. 5). The CHTR response was the exception, where lactate elevation was delayed to the 0.5-hour interval in this treatment. Lactate levels were generally elevated during the first 2 hours of exposure compared to pre-treatment levels. Recovery appeared to begin 2 hours post-treatment for wild fish (Fig. 5) and both wild and cultured fish lactate levels dropped at 24 hours. Though elevation and recovery of lactate was observed, none of the elevation was determined significant.
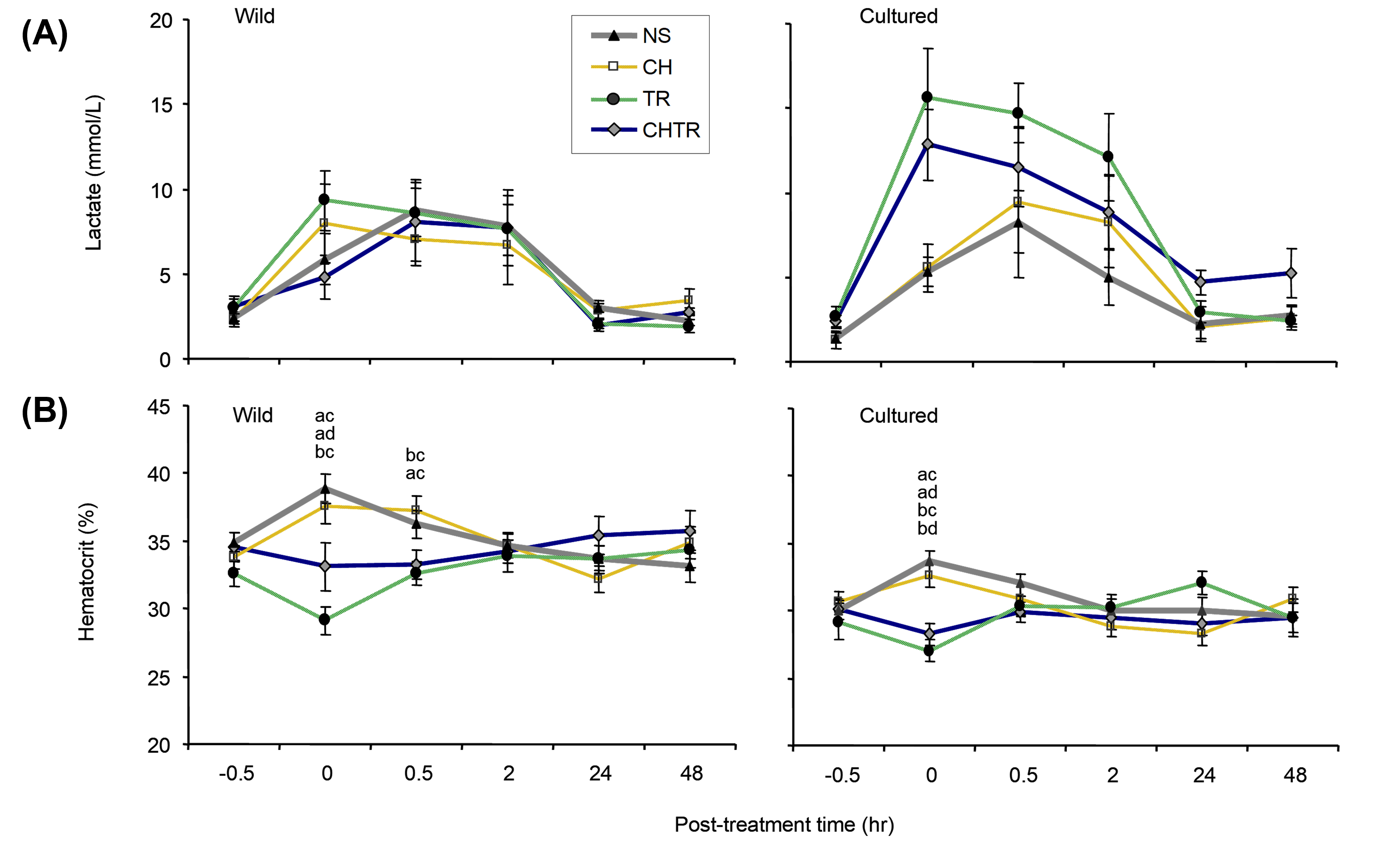
Hematocrit.—The most-marked change in percent hematocrit levels occurred in wild Delta Smelt at the 0-hour interval compared to pre-treatment control conditions (Fig. 5). At this time interval, relative differences between the mean NS and T+R levels were at their greatest. Percent hematocrit readings were essentially similar to the control condition after 2 hours for all treatments and among treatments.
Comparison of Wild and Cultured Smelt Responses
Plasma cortisol, glucose, and lactate responses from wild and cultured Delta Smelt differed markedly. Generally, wild Delta Smelt cortisol levels were higher than cultured Delta Smelt cortisol levels (Figs. 2 and 3). However wild Delta Smelt started with slightly higher and more-variable pre-treatment cortisol levels than cultured Delta Smelt though the difference was not significant.
Cortisol.—On average, cortisol levels in wild and cultured fish were elevated from their baseline levels at 0, 0.5 and 2 hours for all treatments (Fig. 2, P < 0.05). In contrast to wild smelt, cortisol levels for cultured Delta Smelt in the NS, C+H, and T+R were not statistically different from pre-treatment levels by 24-hour post-treatment. C+H and CHTR wild and cultured Delta Smelt cortisol levels were similar immediately after the treatment and then differed within 2 hours shortly afterwards (Fig. 3). Both wild and cultured Delta Smelt in the T+R treatment had similar cortisol levels at all time intervals. In contrast, cortisol levels in the NS experiments were significantly different at all treatment time intervals between cultured and wild fish, except at 48 hours. At 24 hours, both wild and cultured Delta Smelt in the CHTR replicates had higher cortisol levels than wild and cultured smelt in the C+H replicates. Mean cortisol levels tended to increase again after 24 hours for all groups except for wild smelt from the CHTR experiments (Figs. 2 and 3, P < 0.05).
Glucose.—Control condition glucose levels of wild and cultured fish averaged around 64 mg/dL and did not increase or decrease from a pre-treatment level rapidly for any treatment; and glucose elevation follows the elevation of primary stress indicators (the stress hormones, the glucocorticoids and cortisol) Glucose levels in each of the treatments showed no significant differences for wild and cultured fish until 2- and/or 24-hour post-treatment intervals (Fig. 4, P < 0.05).
Glucose levels from wild Delta Smelt were elevated at 2- and 24-hour intervals compared to previous intervals and to cultured smelt (Fig. 4, P < 0.05). In contrast to wild Delta Smelt, cultured Delta Smelt glucose levels remained at levels similar to control condition levels at 0- and 2-hours post-treatment except for cultured smelt in the C+H experiments at 2 hours where there was a slight uptick (Fig. 4). At 48 hours, wild Delta Smelt glucose levels went down whereas cultured Delta Smelt glucose levels showed some fluctuations in elevation at 24-48 hours among all the test groups (Fig. 4). Wild smelt glucose levels were significantly higher at 24 hours in the C+H experiments.
Lactate.—Generally, lactate control condition levels in wild and cultured fish were approximately 2 mmol/L. Lactate levels were elevated at 0 through 2 hours after the experiment replicates but returned to control condition levels at 24 hours. Mean lactate levels for cultured fish from the C+H and NS treatments peaked higher at 0-hour compared to the delayed (0.5 hours) and lower peak concentrations were observed for the T+R and CHTR treatments (Fig. 5).
The lactate response of wild and cultured Delta Smelt showed some marked differences to those observed for cortisol and glucose. The lactate levels for wild fish showed a relatively consistent pattern among the treatments and were very similar to the NS results for cultured fish. The most-noted relative differences were the elevated mean lactate concentrations for cultured smelt at 0-hour for CHTR and at 0- to 2-hours for T+R experiments (Fig. 6). In addition, wild Delta Smelt appeared to have smaller standard errors among its treatment means (i.e., responses appeared less variable) than those of cultured Delta Smelt (Fig. 5).
Hematocrit.—Mean hematocrit levels of wild Delta Smelt were higher than cultured Delta Smelt hematocrit levels at every time interval in the NS, C+H and CHTR treatments (Fig. 7). However, wild and cultured Delta Smelt in the T+R treatments did not have significantly different hematocrit levels at 0-, 0.5-, and 24-hours post-treatment.
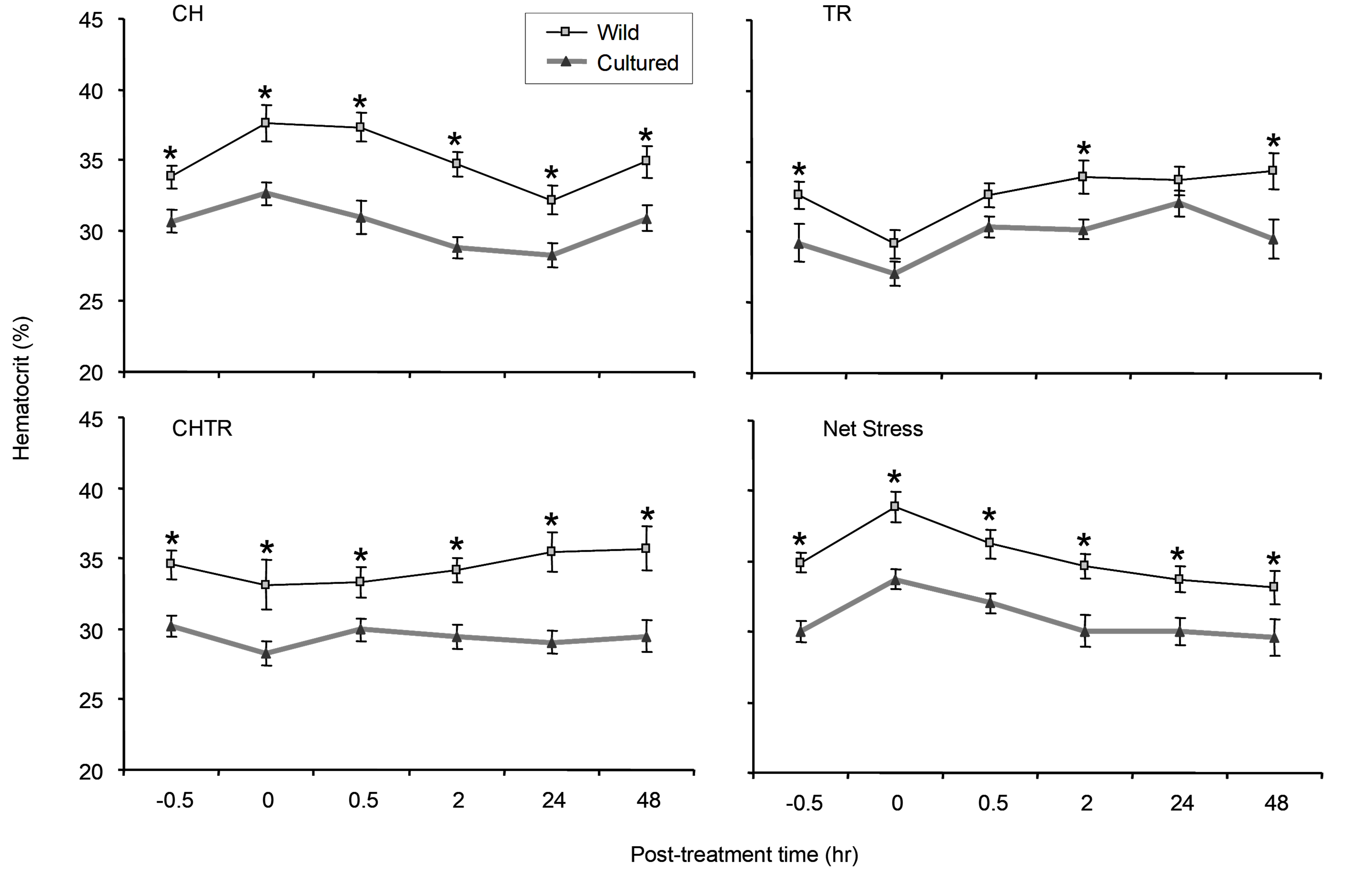
Hematocrit levels were not significantly different from control condition levels throughout the time intervals for most treatments for both cultured and wild fish. Wild smelt showed significant differences at the 0-hour interval for NS, C+H, and T+R and at 0.5 hour for C+H replicates (P < 0.05). Conversely, significant differences were only observed between the control condition and 0-hour interval responses for NS.
Results differed significantly for almost all comparisons between the treatment groups T+R at the 0-hour interval for both wild and cultured (Fig. 5). The exception was the C+H and CHTR at 0-hour for wild fish. In addition, C+H and NS treatments showed a significant difference from T+R treatments at the 0.5-hour interval for wild smelt.
Quality Control Performance Results
Fish and instrument measurements.—Percent deviations from replicate fork length and fish weight measurements were within acceptable limits. Percent deviation for length measurements were between individuals measuring the fish and weight were between repeated measurements on the same scale. Mean percent deviation for fish length measurements was 0.60 ± 1.47%, and all but 1 value was under 5% deviation. Fish weight measurements had a mean percent deviation of 0.41 ± 0.42% and all values were within acceptable levels.
Hematocrit and water quality parameter measurements showed higher levels of measurement variability. Hematocrit quality control readings had a mean percent deviation of 1.57 ± 2.57%. Four hematocrit values out of 157 measurements exceeded 5% deviation. The mean percent deviations for electrical conductivity, dissolved oxygen (mg/L), and water temperature were 2.87, 3.87, and 1.80%. A higher proportion of these measurements exceeded 5% deviation. Some of the large differences in duplicate field measurements for electrical conductivity and dissolved oxygen were attributable to changing conditions in the quality of water held in the tanker truck. The varying measurements may have reflected the slow dissolving and mixing of salt and with the continuous injection of oxygen into the water.
Cortisol analyses.—Cortisol detection sensitivity was 0.30 ± 0.046 pg/well (picograms per microtiter well), or 99.0 ± 15.2 pg/ml (0.099 ± 0.015 ng/ml) with a 50% displacement of enzyme label at approximately 30 to 40 pg/well and a linear response of the standard curve through 250 pg/well. Serial dilutions of selected serum samples produced displacement curves parallel to that of the cortisol standard dose-response curve.
These extracted serum sample samples, diluted for the parallelism study and calculated against the cortisol standard curve, gave recovery values of 97.25 ± 5.49%. The precision of the enzyme immunoassay (EIA) was assessed in selected pooled serum samples. The intra-plate C-V (control values) for cortisol concentrations averaged 3.5 to 5.4% (n = 3) when 20 aliquots of the same sample were assayed in the same plate. The average inter-plate (intra-assay) C-Vs for polls of high, medium, and low cortisol concentrations were 4.1, 7.2, and 10.5%, whereas the average inter-assay C-Vs for these same pools were 7.1, 9.4, and 13.5%.
Data entry accuracy.—We randomly checked 5% of the data for accuracy. An error rate of approximately 25% was discovered triggering corrective action. The corrective action entailed a second line-by-line edit of 100% of the data entered into the database. This editing resulted in a near 0% data entry error rate.
Discussion
General Response and Recovery
For three out of the four physiological parameters examined, adult Delta Smelt physiological stress responses in these experiments showed changes and recovery patterns typical of teleost fishes (Mommsen et al. 1999). Delta Smelt plasma cortisol elevation curve-patterns were similar in shape to those of other teleost fishes, including juvenile Chinook Salmon (Barton et al. 1986; Portz 2007), Lingcod, Ophiodon elongatus (Milston et al. 2006), and Olive Flounder, Paralichthys olivaceus (Hur et al. 2007). Much of the available primary and secondary stress information from fish research has been collected on salmonids. Osmerids and Salmonids are closely related families (Schmidtke and Kandt 1981). However, our experiments indicate Delta Smelt have a more-pronounced elevation of plasma cortisol, glucose, and lactate, peaking higher, and earlier, when compared to salmonids. Similar results were observed by other studies, where juvenile Delta Smelt held at 21 °C demonstrated increased stress response and a reduction in available energy compared to fish held at 17 °C (Pasparakis et al. 2022). Since the magnitude of stress hormones varies widely among species and individuals (Strange et al. 1978; Wedemeyer et al. 1990), we placed more significance on the similarity of response trends described in the literature than on the magnitude of responses.
Cortisol as a Diagnostic Stress Indicator
Wild Delta Smelt showed a greater amplification of cortisol in response to all treatments, including the NS, than cultured Delta Smelt. Wild Delta Smelt cortisol levels were still higher than cultured fish at 24 hours, except from the T+R experiments. Relatively lower mean response for the T+R experiments may be the result of operational routine salt additions during truck transport. Salt addition during transport helps mediate the stress response (Swanson et al. 1996; Urbinati and Carneiro 2006). Reduction in cortisol during barge transport has also been observed with juvenile Chinook salmon and rainbow trout (Congleton et al. 2000).
Both cultured and wild Delta Smelt had significantly elevated cortisol relative to control condition levels for almost all treatments up to 2 hours. In addition, wild fish cortisol levels in the CHTR treatments were significantly elevated for up to 24 hours. These results suggest that after sequential stressors, Delta Smelt failed to return to baseline levels within 24 hours. Prolonged elevation of stress hormones was also observed for other fishes (Milston et al. 2006; Hur et al. 2007). This prolonged elevation of cortisol was contrasted by the response of wild Delta Smelt exposed for 2 hours to a simulated fish screen and varying water flows where peak cortisol levels declined after 30 minutes (Swanson et al. 2001). Delayed recovery may negatively affect their ability to cope with any additional “insults” to homeostasis and may decrease immune resistance to pathogens (Maule and Schreck 1990; Wang et al. 2005).
The full sequence of the CHTR showed the greatest elevation of cortisol and the least amount of recovery during post-experimental period. We attribute this response pattern to the maximum exposure to the stressful events of CHTR processes and experimental handling. This pattern was consistent with current physiological knowledge that prolonged exposure to multiple stressors can produce measurable cumulative effects.
The extent of the CHTR operations on the elevation and recovery of plasma cortisol is unclear. The net-stress treatments showed response trends slightly less or comparable to the other CHTR treatments. The similarities in cortisol response suggest that experimental handling stress may limit our ability to interpret the impacts of CHTR on sublethal stress as measured by cortisol. Swanson et al. (2001) found that wild Delta Smelt experienced significantly higher cortisol concentrations 0.5 hours after experimental handling and release of test fish into the testing apparatus and 0.5 hours after removal and attributed these peaks largely to experimental handling stress. Swanson et al. (2001) found that peak cortisol elevation was additive with increasing flow regimes.
More effort to minimize the stress of sampling could aid in teasing apart actual facility effects. Treatment groups were more difficult to retrieve from the experiments than the undisturbed –0.5-hour fish. Pre-treatment control cortisol values were low because the fish were quickly removed from tanks with hundreds of fish, so less than 30 seconds of netting was required to capture 4 or more undisturbed fish. Retrieving 24 experiment fish from a larger volume of water took multiple people working together and required some amount of perturbation. Even though we did this as quickly as possible, and extracted blood as quickly as possible, some extra handling and time was practically unavoidable with the tools at our disposal. It is expected with a much larger sample size that this effect could be minimized and that greater or more nuanced differences between the CHTR experiments would be observed.
At 48 hours, Delta Smelt cortisol levels for most of the experiments increased and were significantly different from control state except for C+H exposed cultured fish. This response suggests a post-experimental stressor was inhibiting recovery. We hypothesize this response was an artifact due to either prolonged confinement in the smaller 83 L test bins or due to possible disturbance of test fish by repeated sampling from the same bin or tank for the 2-, 24-, and 48-hour samples. This was despite the fact that the 83L bins were used to keep groups of 4 fish separate that were designated to be sampled at the different time intervals. Swanson et al. (2001) also reported slightly elevated cortisol levels in wild Delta Smelt held for 48 hours in separate 5 L containers following exposure to a fish treadmill. Young and Cech (1993) also found that serial sampling of juvenile striped bass caused elevations in cortisol in that species. The factors contributing to the failure to recover could be delayed mortality, density dependence, confinement stress, disturbance, or all the above.
Glucose, Hematocrit, and Lactate: Secondary Indicators
Glucose does not respond as quickly as a primary stress indicator of hyperglycemia and is considered an indicator of longer-term effects of stress and energetic cost (Hattingh 1977; Barton et al. 1988). Glucose concentrations were not significantly elevated between replicates and were only significantly differentiated between wild and cultured fish at 2 hours for CHTR, T+R, NS and at 24 hours for C+H. Wild Delta Smelt in the CHTR phase displayed glucose concentrations that were similar to cultured fish by 48 hours. Although non-significant, cultured Delta Smelt exposed to CHTR also showed greater elevation of glucose at 24 hours compared to other treatments. In general, Delta Smelt experienced glucose elevation up to 24 hours and wild fish responded greater to the treatments versus cultured fish. Conversely, Swanson et al. (2001) reported that juvenile Chinook salmon stressed in a fish treadmill device exhibited hyperglycemia at 0.5-hour post-treatment and recovered by 24 hours.
Hematocrit elevation for the C+H and NS treatments suggests that fish experienced swelling of the red blood cells due to the extrusion of ions, water uptake, and extracellular acidosis (Nikinmaa and Huestis 1984; Nikinmaa et al. 1987) in the C+H and NS replicates. Swanson et al. (2001) reported significantly elevated hematocrit readings of wild Delta Smelt immediately after release into their test apparatus due to erythrocytic swelling and/or erythrocyte addition from splenic contraction (Pearson and Stevens 1991) due to pre-experimental fish handling. The observed hematocrit values decreased slightly but not significantly for the CHTR and T+R, both at 0- and 0.5-hours. We attribute this decrease to the therapeutic effect of salt addition to the truck water in the CHTR and T+R treatments. Similar responses on blood hematocrit and volume were observed from salmon exposed to salinity of 11 ppt (Redding and Schreck 1983). Again, wild fish exhibited this response to a greater degree than cultured fish. The consistently higher hematocrit levels for wild smelt compared to hatchery fish suggest pre-existing physiological differences between these fish groups.
Despite the limitations of the lactate meter, the lactate results contrasted with those of the cortisol and glucose results. Lactate was lower for wild fish in all treatments at all time periods, except the control period and the NS experiments where they were virtually the same for both wild and cultured fish. The peak lactate interval at 0- or 0.5-hours was comparable to the lactate responses of juvenile Chinook salmon exposed to a simulated fish screen and varying flows (Swanson et al. 2001). Increased lactate levels indicate lower muscle fiber performance (Farrell et al. 1979) suggesting wild Delta Smelt had greater muscle conditioning than cultured smelt.
Wild and Cultured Smelt Differences
The stress responses of wild and cultured Delta Smelt differed significantly for 3 out of 4 physiological parameters examined. Cultured Delta Smelt might be more tolerant to anthropogenic stressors or they may be chronically stressed and thus have a muted stress response. Congleton et al. (2000) reported such differences between hatchery and wild Chinook salmon responding to stress from human impacts. Salmonids have been found to be chronically stressed by confinement (Iwama et al. 1997) and confined Delta Smelt may be similarly stressed. Woodward and Strange (1987) suggested that fish reared in hatchery situations are subjected to an unnatural selection pressure that favors individuals with muted stress responses. We propose that the amplified stress responses of the wild Delta Smelt in our study is due to these fish not being habituated to confinement and/or to other conditions typical to fish rearing. We suggest our observed wild Delta Smelt plasma cortisol and glucose mobilization responses may be a more adaptive and representative response to CHTR stressors under actual salvage operations. The relative differences in stress indicators of wild and cultured Delta Smelt also suggest lines of research to improve culturing methods and fish health (e.g., optimum holding capacity) for rearing Delta Smelt.
Conclusions and Recommendations
We demonstrated physiological parameters can be used to assess the stress responses of adult Delta Smelt within an operational fish salvage facility. Delta Smelt undergoing the full CHTR process displayed the most pronounced stress responses. These responses were consistent with the concept that consecutive and prolonged stressors produce greater levels of stress. The extent of experimental stress observed in our study indicates refinements in the fish handling and post-experimental holding conditions are needed to determine the biological significance of stress experienced in the CHTR process. In addition, Swanson et al. (2005) suggest that the measurement of multiple integrated responses is necessary to fully interpret the physiological impacts of anthropogenic stressors on Delta Smelt.
Milston et al. (2006) concluded that fish should not be exposed to successive stressors for at least 24 hours after the initial stressor. A stress-reduction strategy would allow salvaged fish to recover prior to release. Pre-release acclimation may be an effective way to separate sequential stressors and allow for recovery before the final phase of CHTR. For example, salvaged fish could be placed in commercial net-pens to provide a recovery period prior to release.
The stress response observed during the transport phase supports the use of salt and other water conditioners to protect the slime coat of fish and prevent osmotic imbalance as recommended by Swanson et al. (1996). Practical protocols should be developed to allow facility personnel to closely match the salt concentrations of the transport water to plasma concentrations of Delta Smelt and other salvaged fish (Redding and Schreck 1983). Written standard operating procedures for fish handling, enhanced operations staff training, reducing fish densities in the bucket and truck, and releasing fish at night would in combination help to reduce stress and mortality of salvaged fish.
Acknowledgements
This project was funded by the CALFED Conveyance Program (CALFED Agreement #4600002115). We would like to thank the following groups and individuals for their assistance in the planning, preparation, implementation, analysis, and report editing, which made the completion of this project possible: the California Department of Fish and Wildlife: Robert Fujimura, Geir Aasen, Pat Coulston, Dan Odenweller, Bryan Crouch, Jason Dubois, Jamie Gannon, Mike Garcia, Russell Gartz, Justin Graham, Talia Rose Gruen, Caley Hirsch, Michael Holt, Roxanne Kessler, Jennifer Messineo, Jerry Morinaka, Nancy Rosulek Kramer, Jeff Sanchez, Ramiro Soto, Derek Stein, and Robert Vincik; the Department of Water Resources: Jim Odom and the Skinner Fish Facility staff; Don Kurasaka and Division of Engineering; the University of California, Davis (UCD), Smelt Culture Facility: Joan Lindberg and Luke Ellison; the UCD Clinical Endocrinology Laboratory: Coralie Munro and Alejandro Esteller-Vico; the UCD Wildlife, Fish, and Conservation Biology Department: Joseph J. Cech, Christina Swanson, Paciencia Young, Stephanie Chun, Nicole West, Teresa MacColl, and Trilia Chen; the U.S. Bureau of Reclamation: Mark Bowen, Brent Bridges, Bradd Baskerville-Bridges, Don Portz and Zak Sutphin.
Literature Cited
- Aasen, G. A. 1999. Juvenile Delta Smelt use of shallow-water and channel habitats in California’s Sacramento–San Joaquin Estuary. California Fish and Game 85:161–169.
- Adams, S. M., editor. 1990. Biological Indicators of Stress in Fish. Symposium 8. American Fisheries Society, Bethesda, MD, USA.
- Baskerville-Bridges, B., J. C. Lindberg, and J. J. Cech, Jr. 2005. Manual for the intensive culture of Delta Smelt (Hypomesus transpacificus). Report to CALFED Bay–Delta Program, ERP-02-P31, University of California, Davis, CA, USA.
- Baskerville-Bridges, B., J. C. Lindberg, and J. J. Cech, Jr. 2006. Delta Smelt culture, production, and facility expansion, 2003–2005. Calfed Bay-Delta Agreement Number 4600002963. Department of Water Resources, Sacramento, CA, USA.
- Barton, B. A. 2002. Stress in fishes: a diversity of responses with particular reference to changes in circulating corticosteroids. Integrative and Comparative Biology 42:517–525.
- Barton, B. A., C. B. Schreck, and L. G. Fowler. 1988. Fasting and diet content affect stress-induced changes in plasma glucose and cortisol in juvenile Chinook Salmon. Progressive Fish Culturist 50:16–22.
- Barton, B. A., C. B. Schreck, and L. A. Sigismondi. 1986. Multiple acute disturbances evoke cumulative physiological stress responses in juvenile Chinook Salmon. Transactions of the American Fisheries Society 115:245–251.
- Carragher, J. F., and C. M. Rees. 1994. Primary and secondary stress responses in golden perch, Macquaria ambigua. Comparative Biochemistry and Physiology 107(1):49–56.
- Congleton, J. L., W. J. LaVoie, C. B. Schreck, and L. E. Davis. 2000. Stress indices in migrating juvenile Chinook Salmon and Steelhead of wild and hatchery origin before and after barge transportation. Transactions of the American Fisheries Society 129:946–961.
- Farrell, P. A., J. H. Wilmore, E. F. Coyle, J. E. Billing, and D. L. Costill. 1979. Plasma lactate accumulation and distance running performance. Medicine and Science in Sports 11:338–344.
- Hattingh, J. 1977. Blood sugar as an indicator of stress in the freshwater fish, Labeo capensis (Smith). Journal of Fish Biology 10(2):191–195.
- Herrgesell, P. L. 2012. A historical perspective of the Interagency Ecological Program: Bridging multi-agency studies into ecological understanding of the Sacramento-San Joaquin Delta and estuary for 40 Years. Interagency Ecological Program Final Report, California Department of Fish and Game, Sacramento, CA, USA.
- Hur, J. W., I-S. Park, , and J. Y. Chang. 2007. Physiological responses of the olive flounder, Paralichthys olivaceus, to a series stress during the transportation process. Ichthyological Research 54(1):32–37.
- Iwama, G. K., A. D. Pickering, J. P. Sumpter, and C. B. Schreck, editors. 1997. Fish Stress and Health in Aquaculture. Society for Experimental Biology Seminar Series, Volume 62. Cambridge University Press, Cambridge, UK.
- Maule, A. G., and C. B. Schreck. 1990. Changes in numbers of leukocytes in immune organs of juvenile Coho Salmon after acute stress or cortisol treatment. Journal of Aquatic Animal Health 2:298–304.
- McAllister, D. E. 1963. A Revision of the Smelt Family, Osmeridae. National Museum of Canada Bulletin 191. Department of Northern Affairs and National Resources, Ottawa, Ontario, Canada.
- Milston, R. H., M. W. Davis, S. J. Parker, B. L. Olla, S. Clements, and C. B. Schreck. 2006. Characterization of the physiological stress response in Lingcod. Transactions of the American Fisheries Society 135:1165–1174.
- Mommsen, T. P., M. M. Vijayan, and T. W. Moon. 1999. Cortisol in teleosts: dynamics, mechanisms of action, and metabolic regulation. Reviews in Fish Biology and Fisheries 9(3):211–268.
- Morinaka, J. 2013. A history of the operational and structural changes to the John E. Skinner Delta Fish Protective Facility from 1968 to 2010. Interagency Ecological Program for the San Francisco Bay/Delta Estuary, Sacramento, CA, USA.
- Moyle, P. B. 2002. Inland Fishes of California. Revised and expanded. University of California Press, Berkeley, CA, USA.
- Munro, C., and G. Stabenfeldt. 1985. Development of a cortisol enzyme immunoassay in plasma. Clinical Chemistry 31:956.
- Nikinmaa, M., and W. H. Huestis. 1984. Adrenergic swelling of nucleated erythrocytes: cellular mechanisms in a bird, domestic goose, and two teleosts, Striped Bass and Rainbow Trout. Journal of Experimental Biology 113(1):215–224.
- Nikinmaa, M., J. F. Steffensen, B. L. Tufts, and D. J. Randall. 1987. Control of red cell volume and pH in trout: effects of isoproterenol, transport inhibitors, and pH in bicarbonate/carbon dioxide-buffered media. Journal of Experimental Zoology 242(3):273–281.
- Pasparakis, C., A. N. Wampler, T. Lohroff, F. DeCastro, D. E. Cocherell, E. W. Carson, T. C. Hung, R. E. Connon, N. A. Fangue, and A. E. Todgham. Characterizing the stress response in juvenile Delta Smelt exposed to multiple stressors. Comparative Biochemistry and Physiology, Part A Molecular and Integrative Physiology 274:111303. doi.org/10.1016/j.cbpa.2022.111303
- Pearson, M. P., and E. D. Stevens. 1991. Size and hematological impact of the splenic erythrocyte reservoir in rainbow trout, Oncorhynchus mykiss. Fish Physiology and Biochemistry 9(1):39–50.
- Portz, D. E., C. M. Woodley, and J. J. Cech. 2006. Stress-associated impacts of short-term holding on fishes. Reviews in Fish Biology and Fisheries 16(2):125–170.
- Portz, D. E. 2007. Fish-holding-associated Stress in Sacramento River Chinook Salmon (Oncorhynchus tshawytscha) at South Delta Fish Salvage Operations: effects on plasma constituents, swimming performance, and predator avoidance. Dissertation, University of California, Davis, CA, USA.
- Redding, J. M., and C. B. Schreck. 1983. Influence of ambient salinity on osmoregulation and cortisol concentration in yearling Coho Salmon during stress. Transactions of the American Fisheries Society 112(6):800–807.
- Rodbard, D., and J. E. Lewald. 1970. Computer analysis of radioligand assay and radioimmunoassay data. Acta Endocrinology Supplement (Copenhagen) 147:79–103.
- Sapolsky, R. M., L. M. Romero, and A. U. Munck. 2000. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocrine Reviews 21(1):55–89.
- Schmidtke, J., and I. Kandt. 1981. Single-copy DNA relationships between diploid and tetraploid teleostean fish species. Chromosoma (Berlin) 83:191–197.
- Schreck, C. B. 1981. Stress and compensation in teleostean fishes: response to social and physical factors. Pages 295–321 in A. D. Pickering, editor. Stress in Fish. Academic Press, London, UK.
- Schreck, C. B. 1982. Stress and rearing of salmonids. Aquaculture 28:241–249.
- Schreck, C. B., W. Contreras-Sanchez, and M. S. Fitzpatrick. 2001. Effects of stress on fish reproduction, gamete quality, and progeny. Aquaculture 197:3–24.
- Selye, H. 1950. Stress and the general adaptation syndrome. British Medical Journal 1:1383–1392.
- Shanghavi, D. S., and J. M. Weber. 1999. Effects of sustained swimming on hepatic glucose production of rainbow trout. Journal of Experimental Biology 202(16):2161–2166.
- Skinner, J. E. 1974. A functional evaluation of a large louver screen installation and fish facilities research on California water diversion projects. Pages 225–249 in L. D. Jensen, editor. Proceedings of the Second Entrainment and Intake-Screening Workshop. Cooling Water Research Project. Report number 15. Johns Hopkins University, Baltimore, MD, USA.
- Strange, R. J., C. B. Schreck, and R. D. Ewing. 1978. Cortisol concentrations in confined juvenile Chinook Salmon (Oncorhynchus tshawytscha). Transactions of the American Fisheries Society 107(6):812–819.
- Swanson, C., and J. J. Cech. 1996. Environmental tolerances and habitat requirements of Delta Smelt. Final Report to California Department of Water Resources, Sacramento, CA, USA.
- Swanson, C., R. C. Mager, S. I. Doroshov, and J. J. Cech, Jr. 1996. Use of salts, anesthetics, and polymers to minimize handling and transport mortality in Delta Smelt. Transactions of the American Fisheries Society 125:326–329.
- Swanson, C., T. Reid, P. S. Young, and J. J. Cech, Jr. 2000. Comparative environmental tolerances of threatened delta smelt (Hypomesus transpacificus) and introduced wakasagi (H. nipponensis) in an altered California estuary. Oecologia 123(3):384–390.
- Swanson, C., P. S. Young, and J. J. Cech, Jr. 2005. Close encounters with a fish screen: integrating physiological and behavioral results to protect endangered species in exploited ecosystems. Transactions of the American Fisheries Society 134:1111–1123.
- Swanson, C., P. S. Young, S. Chun, T. Chen, T. MacColl, L. Kanemoto, and J. J. Cech, Jr. 2001. Fish treadmill-developed fish screen criteria for native Sacramento-San Joaquin watershed fishes. Part 2 Biological Studies. Final Report to CALFED, California Department of Water Resources, Sacramento, CA, USA.
- Urbinati, E. C., and P. C. F. Carneiro. 2006. Sodium chloride added to transport water and physiological responses of Matrinxa, Brycon amazonicus (Teleost: Characidae). Acta Amazonica 36(4):569–572.
- Young, P. S., and J. J. Cech, Jr. 1993. Physiological stress responses to serial sampling and confinement in young-of-the-year striped bass, Morone saxatilis (Walbaum). Comparative Biochemistry and Physiology, Part A: Molecular and Integrative Physiology 105A(2):239–244.
- Young, P. S., C. Swanson, S. Chun, T. Chen, T. MacColl, and J. J. Cech, Jr. 2001. Responses and recovery in Delta Smelt exposed to handling stress during fish treadmill experiments at winter temperature. Interagency Ecological Program for the Sacramento-San Joaquin Estuary. IEP Newsletter 14(3):38–40.
- Wang, W. B., A. H. Li, T. Z. Cai, and J. G. Wang. 2005. Effects of intraperitoneal injection of cortisol on non-specific immune functions of Ctenopharyngodon idella.
Journal of Fish Biology 67(3):779–793. - Wedemeyer, G. A. 1996. Physiology of fish in intensive culture systems. Chapman & Hall, New York, NY, USA.
- Wedemeyer, G. A., B. A. Barton, and D. J. McLeay. 1990. Stress and acclimation. Pages 451–477 in C. B. Schreck and P. B. Moyle, editors. Methods for Fish Biology. American Fisheries Society, Bethesda, MD, USA.
- Wendelaar Bonga, S. E. 1997. The stress response in fish. Physiological Reviews 77(3):591–625.
- Woodward, C. C., and R. J. Strange. 1987. Physiological responses in wild and hatchery-reared rainbow trout. Transactions of the American Fisheries Society 116:574–579.
- Zar, J. H. 1999. Biostatistical Analysis. 4th edition. Prentice-Hall, Upper Saddle River, NJ, USA.